Documentos de Académico
Documentos de Profesional
Documentos de Cultura
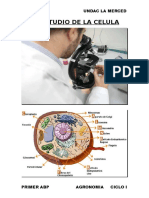
Unidad 8 La Célula Eucariota
Cargado por
marcosgarciad06Derechos de autor
Formatos disponibles
Compartir este documento
Compartir o incrustar documentos
¿Le pareció útil este documento?
¿Este contenido es inapropiado?
Denunciar este documentoCopyright:
Formatos disponibles
Unidad 8 La Célula Eucariota
Cargado por
marcosgarciad06Copyright:
Formatos disponibles
1
ESTRUCTURA CELULAR. LA CÉLULA EUCARIOTA: ESTRUCTURA Y FUNCIONES
GENERALES DE LOS ORGÁNULOS CELULARES.
1. TEORÍA CELULAR
• En 1665, Robert Hooke observó al microscopio un fragmento de corcho y
descubrió que estaba formado por estructuras parecidas a las celdas de los
panales de las abejas, por los que estas estructuras que observó las llamó células
o cellulas.
• Más tarde en 1838, y gracias al desarrollo de la microscopía Scheleiden y
Schwan (el primero para los vegetales y el segundo para los animales)
enunciaron el principio básico de la teoría celular según el cual todos los tejidos
animales y vegetales están compuestos por una o más células, de manera que la
célula es la unidad básicas de todos los organismos.
• En 1855, Virchow estableció que todas las células provienen de otras células
preexistentes (omnis cellula e cellula).
José Juan Canel Álvarez
2
Actualmente, la Teoría celular se resume en los siguientes puntos:
1. Todos los seres vivos están formados por una o varias células, de manera que
podemos dividir a los seres vivos en dos grandes grupos:
a. Seres Unicelulares: Formados por una célula.
b. Seres Pluricelulares: Formados por más de una célula.
2. La célula es la unidad estructural y fisiológica de los seres vivos.
3. Las células son las unidades básicas de reproducción, ya que toda célula proviene
de la división de otras células preexistentes y es idéntica a éstas genética, estructural y
funcionalmente.
4. La célula es la unidad de vida independiente más elemental.
Además, como excepción podemos decir que los virus no son más que simples
moléculas de ácidos nucleicos, recubiertos, como ya veremos de proteínas y a estos
organismos se les llama organismos acelulares. Son parásitos obligados porque
siempre infectan una célula y no se les considera seres vivos porque cuando están
fuera de la célula hospedadora no pueden realizar ninguna función vital
(nutrición, relación y reproducción).
En 1970, Lynn Margulis, propuso la Teoría endosimbiótica o simbiogénesis, donde
afirma que la simbiosis (asociación entre organismos en la que se benefician
mutuamente) en un mecanismo clave en la evolución. Cuando lleguemos a las
mitocondrias y cloroplastos veremos con más detalle la Teoría endosimbiótica.
. CÉLULAS PROCARIOTAS Y CÉLULAS EUCARIOTAS
1. Células Procariotas: Son muy simples y apenas tienen estructuras en su interior. No
tienen núcleo, es decir, su material genético no está separado del citoplasma. Algunas
ejemplos son las bacterias y las cianofíceas.
2. Células Eucariotas: Son las típicas del resto de los organismos ya sean uni o
pluricelulares, animales o vegetales. Tienen orgánulos celulares y poseen un núcleo que
separa su material genético del citoplasma. Hay dos clases: Animal y vegetal.
3. ESTRUCTURAL GENERAL DE UNA CÉLULA EUCARIOTA
El estudio de la célula y de sus orgánulos o compartimentos celulares, lo dividimos en
las siguientes partes:
1. Membrana Plasmatica.
ì ì ì ìRugoso.
ï ï
ï ï1. Reticulo endoplasmaticoí .
ï ï ï ïLiso.
ï ï î
ï
ï ï ï2. Aparato de Golgi.
ï ï ï
ï ï ï
ï
3. Vacuolas.
ï
ïOrganulos con membrana í4. Lisosomas.
ï
ï1.
ï Citoplasma con organulosí ï
2. Protoplasma í ï ï5. Mitocondrias.
ï ï ï
ï ï ï6. Plastos.
ï ï ï7. Peroxisomas.
ï ï ï
ï ï ï
î
ï ï
ï îEstructuras celulares ( Sin membrana ) : Ribosomas.
ï
ï
î2.
ï Hialoplasma ( citoplasma sin organulos ) : Citoesqueleto.
ì1. Envoltura nuclear.
ï
ï2. Nucleoplasma o jugo nuclear.
3. Nucleo í
ï3. Nucleolo.
ï4. Cromatina.
î
José Juan Canel Álvarez
3
Además, hay que tener en cuenta que los orgánulos según se observen al microscopio
óptico o electrónico presentan un aspecto distinto, por eso hablaremos de una
estructura (microscopio óptico) y una ultraestructura celular (microscopio
electrónico).
4. LAS MEMBRANAS BIOLÓGICAS: LA MEMBRANA UNITARIA.
La membrana plasmática y los orgánulos que hay dentro de la célula están formados por
membranas, y estas membranas son barreras o fronteras que permiten no sólo separar,
sino también poner en comunicación distintos compartimentos en el interior de la
célula, así como a la propia célula con el exterior.
La estructura de todas estas membranas es muy parecida, y las diferencias que hay entre
ellas es más bien a nivel de función particular que tienen los distintos orgánulos
formados por estas membranas; por eso, las distintas funciones que van a tener estos
orgánulos va a depender de la composición en proteínas que tengan sus membranas
biológicas.
Los orgánulos formados por membranas son:
1. MEMBRANA PLASMÁTICA.
2. RETÍCULO ENDOPLASMÁTICO.
3. APARATO DE GOLGI.
4. LISOSOMAS.
5. PEROXISOMAS.
José Juan Canel Álvarez
4
6. MITOCONDRIAS.
7. PLASTOS.
8. VACUOLAS.
9. ENVOLTURA O MEMBRANA NUCLEAR.
• COMPOSICIÓN DE LAS MEMBRANAS
Las membranas de las células eucariotas están formadas por una BICAPA LIPÍDICA
A LA QUE SE UNEN MÁS O MENOS FUERTEMENTE UNA SERIE DE
PROTEÍNAS Y OLIGOSACÁRIDOS (en la membrana plasmática) (Modelo del
Mosaico fluido de Singer y Nicholson), teniendo en cuenta que la membrana
plasmática lleva oligosacáridos que sólo están por la parte de fuera y que forman el
llamado glucocálix.
Dicho estos vamos a pasar a estudiar los distintos componentes de las membranas:
1. Lípidos:
Los lípidos más abundantes son:
a) Fosfolípidos.
b) Colesterol (menos en la membrana interna de las mitocondrias que no tiene
colesterol).
c) Glucolípidos (Por la parte de fuera de la membrana plasmática).
Estos lípidos como ya sabemos debido a su carácter anfipático (recordar que tienen
partes polares y partes no polares) en medio acuoso forman una bicapa; esta bicapa
aporta la estructura básica de la membrana y debido a su fluidez es posible muchas
funciones que realizan las membranas celulares.
La bicapa es fluida porque se comporta igual que lo hace un líquido es decir las
moléculas pueden desplazarse sobre sí mismas intercambiando posiciones, y la fluidez
de las membranas dependen de cuatro factores:
• Cuanto más ácidos grasos saturados, la fluidez disminuye, ya que
disminuye la libertad de movimiento dentro de la bicapa.
• Cuanta mayor sea la longitud de las cadenas de los ácidos grasos, la fluidez
disminuye.
• A menor temperatura, menor fluidez.
• A mayor proporción de colesterol, menos fluidez, ya que el colesterol es una
molécula muy rígida.
2. Proteínas:
La posición que ocupan en la bicapa lipídica va a depender de su mayor o menor
afinidad por el agua, de manera que las partes hidrófilas de la proteína quedan hacia el
interior o exterior de la bicapa lipídica, y las partes hidrófobas se sitúan en el seno de la
bicapa lipídica y gracias a estos distinguimos los siguientes tipos de proteínas:
a) Proteínas Intrínsecas o Integrales: Son proteínas que están unidas de una manera
muy fuerte a los lípidos, y que sólo separan de ellos con detergentes que son capaces de
disolver a los lípidos. Dentro de esta clase de proteínas distinguimos a su vez dos clases:
• Proteínas Transmembranales: Atraviesan la bicapa lipídica totalmente.
• Proteínas Integrales o proteínas monotópicas ya que sólo atraviesan una capa.
Las proteínas integrales son anfipáticas, con su partes polares expuestas al medio
acuoso y las partes hidrófobas insertadas en la bicapa.
José Juan Canel Álvarez
5
b) Proteínas Extrínsecas o Periféricas: Son proteínas que están unidas débilmente a la
bicapa lipídica y se separan de ésta por tratamientos débiles. Como las anteriores
también las podemos dividir en dos clases:
• Proteínas Periféricas asociadas a las proteínas integrales.
• Proteínas Periféricas asociadas a los lípidos de la bicapa.
3. Oligosacáridos:
Pueden estar unidos a los lípidos formando entonces los GLUCOLÍPIDOS o pueden
estar unidos a las proteínas formando las GLUCOPROTEÍNAS. ESTÁN POR LA
CARA EXTERNA DE LA MEMBRANA PLASMÁTICA Y FORMAN LA
CUBIERTA CELULAR LLAMADA GLUCOCÁLIX O GLICOCÁLIX, de
manera que las funciones más importantes del glicocálix son:
1. Reconocimiento celular.
José Juan Canel Álvarez
6
2. Se cree que facilita la orientación de ciertas proteínas contribuyendo así a que no se
tumben en la bicapa de manera que las proteínas mantengan su estructura plegada.
3. Protege a la superficie de la célula de lesiones.
Además, otra cosa importante es que esta disposición de los oligosacáridos sólo por la
cara externa de la membrana plasmática, y el hecho de que los lípidos sean distintos en
las dos partes de la membrana hace que LAS MEMBRANAS SEAN ASIMÉTRICAS,
es decir las membranas son distintas por un lado que por el otro.
Por último, decir que este modelo de membrana y en particular el modelo de membrana
plasmática fue propuesto por Singer y Nicholson en 1972 y se llama MODELO DEL
MOSAICO FLUIDO y sus características más importantes a modo de resumen son:
1. Los lípidos y las proteínas integrales que forman las membranas forman un
mosaico molecular.
2. Los lípidos y las proteínas pueden desplazarse por la bicapa lipídica, es decir
LAS MEMBRANAS SON FLUIDAS.
3. LAS MEMBRANAS SON ASIMÉTRICAS, es decir son distintas por un lado y
por el otro, y esta asimetría implica que las dos caras realicen funciones distintas.
• También hay que decir que para el estudio de la membrana plasmática se emplean
eritrocitos (glóbulos rojos) ya que tienen la ventaja que en su interior no tienen
ningún otro sistema de membranas:
Primero se aíslan los glóbulos rojos de la sangre mediante centrifugación para luego
introducirlos en un medio hipotónico como por ejemplo la agua destilada. Con esto se
logra que entre agua en los eritrocitos (recordar la ósmosis) de manera que estos
hinchan y explotan. Posteriormente se aíslan las membranas plasmáticas del resto de los
componentes por centrifugación y se procede a su estudio.
• Por último, la membrana plasmática vista al microscopio electrónico posee un grosor
aproximado de 75Aº, y se observa una estructura típica de tres bandas, donde las
bandas oscuras son las partes hidrófilas de los lípidos y de las proteínas y las bandas
claras son las cadenas hidrocarbonadas.
José Juan Canel Álvarez
7
FUNCIONES DE LA MEMBRANA PLASMÁTICA
1. Función de Intercambio: La membrana plasmática es básicamente una barrera de
permeabilidad, limita a la célula e impide el paso de sustancias (no de todas, pero sí de
muchas), tanto del exterior al interior, como al revés.
2. Función Receptora: Como por ejemplo algunas proteínas de la membrana
plasmática que son receptores específicos de hormonas, de manera que, al existir
distintos receptores específicos en la membrana plasmática de las células, y al tener las
células distintos receptores específicos, la actividad de cada célula será distinta.
3. Función De Reconocimiento: Esta función se realiza gracias a los oligosacáridos y
a las proteínas de la cara externa de la membrana plasmática. Por ejemplo, las
células del sistema inmunológico que nos defienden de los agentes patógenos van a
reconocer las células que son del propio organismo diferenciándolas de las extrañas
gracias a los oligosacáridos y proteínas de la membrana plasmática.
DIFERENCIACIONES DE LA MEMBRANA PLASMATICA
Las más importantes son las siguientes:
1. Microvellosidades: Las suelen tener las células que por sus funciones necesitan
una gran superficie, como por ejemplo las células del intestino, y las
microvellosidades consisten ni más ni menos en muchos repliegues de manera
que se aumenta la superficie de la membrana plasmática.
2. Uniones intercelulares:
Sólo se dan en células animales y son las uniones entre células vecinas por sus
membranas. Destacaremos las siguientes:
a) Uniones de adherencia: Mantienen unidas a las células vecinas pero las
membranas de las células no se fusionan y hacen que las células funcionen como
una unidad, de forma que se encuentran en tejidos donde hay fuertes tensiones
mecánicas como por ejemplo en el músculo cardíaco o en el cuello del útero. Un
ejemplo de uniones de adherencia son los desmosomas.
José Juan Canel Álvarez
8
b) Uniones Estrechas u Oclusivas o Impermeables: Sellan células vecinas por medio
de proteínas por lo que desparece el espacio entre las dos células e impide el paso de
moléculas por estas uniones. Son típicas de los epitelios.
c) Uniones comunicantes, en hendidura o gap: Es típico de las células animales.
Unen las membranas de células vecinas por canales proteicos de forma que permiten el
paso de moléculas pequeñas y de impulsos eléctricos.
EL TRANSPORTE A TRAVÉS DE LA MEMBRANA
Como ya dijimos es la función más característica de la membrana plasmática, y
podemos distinguir los siguientes tipos de transporte:
Transporte
Transporte de moléculas
grandes envueltas en una
De moléculas pequeñas
membrana(Transporte
citoquímico)
Transporte pasivo Transporte activo
A favor de gradiente de En contra de gradiente de
concentración o eléctrico concentración o eléctrico
Difusión simple Difusión facilitada Bombas
Proteínas Canal Transportadores ABC
Permeasas
José Juan Canel Álvarez
9
TRANSPORTE PASIVO:
Es a favor de gradiente de concentración o eléctrico por lo que no gasta energía
(ATP).
LA DIFUSIÓN SIMPLE ES UN PROCESO NO SELECTIVO QUE NO REQUIERE DEL APORTE
DE ENERGÍA
En la difusión pasiva las moléculas atraviesan la bicapa lipídica. Es no selectiva porque no
interviene ninguna proteína de membrana que facilite el transporte. No requiere del aporte de
energía, porque el flujo de las sustancias transportadas se produce a favor gradiente de
concentración o eléctrico.
LA DIFUSIÓN FACILITADA ES UN PROCESO SELECTIVO QUE NO REQUIERE DEL APORTE DE
ENERGÍA.
Es un proceso selectivo, porque requiere de proteínas integrales de membrana, que interaccionan
específicamente con la sustancia transportada. No requiere del aporte de energía, porque las sustancias
transportadas se mueven a favor de su gradiente químico (concentración) y en el caso de aquellas que
tienen carga, también participa el gradiente eléctrico (es decir en función de la distribución de cargas a
uno y otro de la membrana). La difusión facilitada es utilizada, en general, por moléculas con carga
(ácidos orgánicos, aminoácidos, nucleósidos, iones…)
Existen dos variedades de proteínas integrales transportadoras que intervienen en la difusión facilitada:
1) Las proteínas canal.
2) Proteínas transportadoras o Permeasas.
1) Las proteínas Canal:
Como su nombre indica, forman canales en la bicapa, por lo que son proteínas
transmembranales. Son muy específicas y sólo transportan un tipo de molécula o a
veces moléculas muy relacionadas entre sí.
José Juan Canel Álvarez
10
Algunas proteínas canal, como en el dibujo anterior, están siempre abiertas, pero otras
pueden abrirse o cerrarse como respuesta a una señal eléctrica (canal regulado por
voltaje) o porque se les una molécula (canal regulado por ligando), como vemos en el
dibujo siguiente:
2) Las permeasas:
La interacción de la molécula con su transportador implica un cambio conformacional
del transportador, pero no de la molécula transportada.
Por otra parte, las proteínas transportadoras se pueden clasificar en:
1. Uniportes o Uniportadores.
2. Cotransportadores.
José Juan Canel Álvarez
11
3. TRANSPORTE ACTIVO:
Es en contra gradiente de concentración o eléctrico por lo que gasta energía (ATP).
Destacaremos dos clases:
1. Bombas o Bombas iónicas.
2. Transportadores ABC
1. Bombas:
Un ejemplo es la Bomba Sodio-Potasio: Por cada molécula de ATP que se gasta sacan
tres sodios y meten dos potasios, de manera que así se logra que la concentración de
sodio sea 10 veces mayor en el exterior respecto al interior de la célula, y que la
concentración de potasio sea 10 veces mayor en el interior de la célula que en el
exterior, y esto es la base del impulso nervioso.
Estructura de la Bomba Sodio-Potasio:
• Es una ATPasa con 4 subunidades (dos subunidades alfa y dos subunidades
beta).
• En las subunidades alfa se unen el ATP, los 3 iones sodio (por la parte del
citoplasma), y los 2 iones potasio por fuera de la célula.
• Las subunidades beta tienen la función de mantener a las subunidades alfa
en contacto y dentro de la membrana.
Funcionamiento de la Bomba Sodio-Potasio:
Nota para entender el funcionamiento: Cuando la Bomba lleva unido un P tiene poca
afinidad por el Sodio y mucha por el potasio. Si no lleva unido nada, viceversa.
• Por la parte del citoplasma (interior de la célula) la bomba coge tres iones de
sodio y una molécula de ATP que hidroliza (obtiene energía de la rotura del
enlace de un fosfato, dando ADP y P).
• Esta energía hace que cambie de conformación, desplaza los 3 iones al
exterior de la membrana y los suelta en el exterior de la célula, ya que cuando la
bomba lleva unido un P, tiene muy poca afinidad por el sodio.
• Se unen dos iones de potasio por la parte de fuera de la célula, ya que tiene
mucha afinidad por el potasio.
José Juan Canel Álvarez
12
• Al unirse los dos potasios, se libera el P.
• Sin P tiene poca afinidad por el potasio por lo que los suelta en el interior de
la célula y mucha por el sodio, por lo que vuelve a empezar el proceso.
2. Transportadores ABC
José Juan Canel Álvarez
13
2. TRANSPORTE DE MOLÉCULAS GRANDES (TRANSPORTE DE
SUSTANCIAS ENVUELTAS EN UNA MEMBRANA). TRANSPORTE
CITOQUÍMICO
Se trata de un mecanismo que asegura la nutrición y la salida de productos de desecho
de la célula. Veremos tres procesos:
1. Endocitosis.
2. Exocitosis.
3. Transcitosis.
1. Endocitosis:
Es la entrada en la célula de sustancias envueltas en vesículas formadas a partir de la
membrana plasmática. Existen dos tipos de endocitosis:
• Fagocitosis: Las partículas que se introducen son partículas sólidas grandes. Se
realiza por invaginación de la membrana, y la partícula pasa al citoplasma en
forma de vacuola.
• Pinocitosis: Las sustancias que entran son sustancias líquidas o sólidas, pero de
tamaño pequeño. Estas sustancias al igual que en el caso anterior forman al
atravesar la membrana pequeñas vacuolas las cuales pueden juntarse y formar
vacuolas de mayor tamaño.
2.Exocitosis:
Consiste en la salida o excreción de sustancias por medio de vesículas de exocitosis las
cuales se unen a la membrana plasmática y se abren al exterior expulsando su contenido
fuera de la célula.
Después de realizarse la exocitosis, lógicamente la membrana de la vacuola queda
incluida en la membrana plasmática, por lo tanto, una cosa muy importante es que por la
endocitosis la membrana plasmática se hace más pequeña, pero este descenso de tamaño
de la membrana plasmática se compensa con el aumento que se produce de ésta gracias
a la exocitosis.
3.Transcitosis:
Es el conjunto de fenómenos que permiten a una sustancia travesar todo el citoplasma
desde un polo al otro de la célula, sin que la sustancia sufra ninguna transformación.
José Juan Canel Álvarez
14
¿QUÉ OCURRE DESPUÉS DE LA ENDOCITOSIS? LA HETEROFAGIA Y LA
AUTOFAGIA.
HETEROFAGIA:
Después de que ocurre la endocitosis se forma una vesícula o una vacuola en el interior
de la célula que se llama FAGOSOMA. Este fagosoma se une a los Lisosomas
Primarios (son lisosomas que no han actuado nunca) y se forma una vacuola mayor que
se llama FAGOLISOSOMA. En el fagolisosoma las grandes moléculas (polisacáridos,
ácidos nucleicos, proteínas…) se convierten en moléculas más pequeñas
(monosacáridos, bases nitrogenadas, aminoácidos, etc.) ya que como veremos más
adelante los lisosomas tienen en su interior unos enzimas que hidrolizan (= rompen) las
moléculas grandes y las transforman en moléculas más pequeñas.
Después estas partículas pequeñas pasan a través del interior del fagolisosoma al
citoplasma de la célula, quedando en el fagolisosoma las moléculas que no se han
degradado, y este fagolisosoma ahora recibe el nombre de lisosoma secundario
(lisosoma que ya ha actuado y que puede unirse a otros fagosomas). Este lisosoma
secundario como acabamos de decir puede unirse a otros fagosomas de manera que las
sustancias no degradadas se van acumulando progresivamente en estos lisosomas
secundarios.
En ciertos organismos estos lisosomas secundarios pueden fusionarse con la membrana
plasmática y mediante exocitosis expulsan su contenido fuera de la célula, en cambio en
las células de los organismos pluricelulares, lo más normal es que los lisosomas
secundarios se transformen en cuerpos residuales de manera que cuantos más cuerpos
residuales posea una célula, más vieja es.
AUTOFAGIA:
Se produce cuando la digestión es de orgánulos propios de la célula. La autofagia es
importante para las células, ya que destruye zonas dañadas, interviene en el desarrollo
celular y permite la nutrición de la célula cuando las condiciones son desfavorables.
José Juan Canel Álvarez
15
5. FLUJO DE MATERIALES DENTRO DE LA CÉLULA
El conjunto de orgánulos que realizan esta función forma los sistemas de membranas
del citoplasma de la célula, y estos orgánulos son:
1. Retículo Endoplasmático o Endoplásmico.
2. Aparato de Golgi.
3. Lisosomas.
4. Vacuolas.
RETÍCULO ENDOPLASMÁTICO
Tiene una membrana más delgada que la membrana plasmática, pero tiene una
composición similar. El retículo endoplasmático es un complejo sistema de tubos, sacos
y cisternas que pueden llegar a ocupar gran parte de la célula, y es importante destacar
que la porción del Retículo Endoplasmático Rugoso (RER) que hace frontera entre el
núcleo y el hialoplasma forma la envoltura nuclear.
Existen dos tipos de Retículo Endoplasmático:
1. Retículo Endoplasmático Rugoso (RER). 2. Retículo Endoplasmático Liso (REL).
• El RER posee ribosomas adheridos a su membrana por la parte de fuera, en
cambio el REL no los posee; además los ribosomas se unen a la membrana
del RER por su subunidad mayor y por unas glicoproteínas
transmembranales del RER que se llaman Translocones.
Como el RER tiene adosado en su exterior ribosomas, es lógico pensar que éste
estará muy desarrollado en células que por su función realicen una gran labor de
síntesis de proteínas como por ejemplo las células del páncreas o las células del
hígado (se sabe que, si por ejemplo se somete a un animal a un ayuno
prolongado, el RER de las células pancreáticas se reduce considerablemente,
pudiendo volver a recuperarse cuando sometemos al animal a una dieta rica).
• El REL es escaso en la mayor parte de las células, pero es abundante por
ejemplo en las células musculares estriadas donde se llama retículo
sarcoplásmico y que es muy importante en la liberación de Ca2+ que participa en
la contracción muscular o el los hepatocitos (células del hígado), donde
interviene en la síntesis de lípidos para su exportación y en la detoxificación de
compuestos.
José Juan Canel Álvarez
16
APARATO DE GOLGI
Posee una membrana con un grosor aproximado de 60-75 Aº, y se localiza entre el
Retículo endoplasmático y la membrana plasmática. El Retículo Endoplasmático y el
Aparato de Golgi (AG) no están en comunicación “física”, pero como vamos a ver hay
vesículas que van del RER al AG, y estas vesículas se llaman Vesículas de Transición,
las cuales se forman a partir de una zona del RER que no tiene ribosomas y que se llama
Retículo de Transición.
El AG está formado por un conjunto se sacos concéntricos muy apretados, y cada
conjunto de sacos recibe el nombre de Dictiosomas pudiendo encontrar en una célula
desde cinco dictiosomas hasta algunas decenas, en función del tipo de célula y de su
estado funcional.
Los Dictiosomas tienen dos caras:
a. Cara de Formación o Cara Cis o Cara Cóncava: Es la cara más próxima al RER y
alrededor de ella se sitúan las Vesículas de Transición que provienen del RER.
b. Cara de Maduración o Cara Trans o Cara Convexa: Es la más alejada del RER y
de ella se desprenden unas pequeñas vesículas que se llaman Vesículas de Secreción.
El AG está continuamente transformándose gracias a estas vesículas ya que a
partir de las vesículas de transición se forma nuevo AG, el cual se destruye para
formar las vesículas de secreción.
Lógicamente el AG está muy desarrollado en células que realizan funciones de
secreción, como por ejemplo las células secretoras del mucus del epitelio intestinal o las
células del tiroides.
José Juan Canel Álvarez
17
LISOSOMAS
Son pequeñas vesículas que se originan a partir de la cara de maduración del AG.
Se caracterizan por tener en su interior enzimas hidrolíticos de manera que rompen las
grandes moléculas dando lugar a otras más pequeñas (acordarse de la heterofagia y de la
autofagia). Estos enzimas hidrolíticos se encuentran en el interior de los lisosomas
porque si estuviesen fuera y libres en el citoplasma de la célula podrían destruir las
estructuras celulares.
Los lisosomas tienen aproximadamente unos 42 enzimas del tipo hidrolasas ácidas (las
hidrolasas ácidas son glicoproteínas y son enzimas cuyo pH óptimo de funcionamiento
es entre 3 y 6, y este pH se mantiene dentro del lisosoma gracias a que éstos poseen en
su membrana una proteína de transporte de protones, que gastando energía (ATP) mete
dentro del lisosoma protones de manera que el interior de los lisosomas se vuelve
ácido).
Podemos distinguir dos clases de lisosomas:
1. Lisosomas Primarios: Son los lisosomas recién formados a partir de los dictiosomas
del AG y que sólo contienen los enzima hidrolíticos.
2. Lisosomas Secundarios: Son los lisosomas que ya han actuado y que por lo tanto
contienen en su interior los enzimas digestivos más sustancias que no se han
descompuesto (recordar la heterofagia y la autofagia).
PEROXISOMAS
• Sólo se encuentran en las células animales y se forman por gemación del RER.
• Son semejantes a los lisosomas, pero no poseen hidrolasas ácidas, pero sí
enzimas oxidativos siendo el más importante la Catalasa o Peroxidasa, el cual es
un enzima que transforma la reacción 2H2O2 ® 2H2O + O2.
José Juan Canel Álvarez
18
• En las semillas en germinación de vegetales existe un tipo especial de
peroxisomas que reciben el nombre de Glioxisomas los cuales transforman los
ácidos grasos de las semillas en glúcidos (mediante una serie de reacciones
químicas que se llama Ciclo del Glioxilato) y que son necesarios para que se
desarrolle el embrión (los Glioxisomas no existen en células animales).
Las funciones más importantes de los peroxisomas son:
1. Detoxificación: Ya que además de la catalasa tienen otros enzimas que eliminan
productos muy tóxicos para la célula como el agua oxigenadas, el etanol…y que
son muy importantes para las células del hígado y del riñón.
2. Acortan los ácidos grasos de cadena larga para que luego se acaban de oxidar
en las mitocondrias.
3. Oxidan aminoácidos.
4. Intervienen en la síntesis de fosfoglicéridos y TAG.
LAS VACUOLAS
Están constituidas por una membrana que se llama tonoplasto. Las células animales
tienen numerosas vacuolas de pequeño tamaño, en cambio las células vegetales
tienen vacuolas de mayor tamaño y son menos numerosas.
En cuanto a la función de las vacuolas son:
1. Almacenan sustancias de reserva y/o
sustancias tóxicas.
2. Almacenan pigmentos como los que les
dan color a los pétalos de las flores.
3. Vacuolas Pulsátiles: Por ejemplo, el
Paramecio es un protozoo que vive en el agua. El
citoplasma del paramecio es hipertónico respecto al
exterior de manera que está continuamente
entrando agua dentro de la célula (acordarse de la
presión osmótica), lo que llevaría a que éste
explotara; entonces las vacuolas pulsátiles lo que
hacen es expulsar el agua que entre dentro del
citoplasma del paramecio, ya que cuando están
llenas se fusionan a la membrana plasmática y
sueltan el agua.
4. Vacuolas Digestivas: Son los fagolisosomas.
FUNCIONES DE LOS SISTEMAS MEMBRANOSOS
• FUNCIONES DEL RER
1) Síntesis de proteínas:
Las proteínas que se fabrican en el RER pueden ser de cuatro tipos:
• Proteínas intrínsecas.
• Proteínas extrínsecas de la cara externa.
• Proteínas de los lisosomas (hidrolasa ácidas).
• Proteínas de secreción.
José Juan Canel Álvarez
19
2) Glicosilación de proteínas: La mayoría de las proteínas sintetizadas en el RE rugoso
son glicosiladas (Glicosilar es unir hidratos de carbono a las proteínas, es decir, es
formar glicoproteínas). La glicosilación se realiza en el lumen del RER, donde un lípido
de la membrana del RER que se llama dolicol transfiere los azúcares hasta la proteína
• FUNCIONES DEL REL
1. Síntesis de Lípidos:
Todos los lípidos de membrana, menos los ácidos grasos (que se sintetizan en el
hialoplasma), se sintetizan en el REL. Por lo tanto, el REL es el lugar donde se
fabrican los componentes de las membranas celulares, los cuales luego son
exportados en vesículas, las cuales se desprenden y se dirigen hacia la membrana de
un orgánulo concreto o hacia la membrana plasmática en la cual se integran. Para
orientar los lípidos en las membranas actúa un enzima que se llama Flipasa, de
forma que la distinta afinidad de la flipasa por los diferentes lípidos es una de las
causas que origina la asimetría de la membrana plasmática.
José Juan Canel Álvarez
20
2. Almacenamiento de Calcio: Gracias a una bomba de Calcio que lo introduce
dentro del REL mediante un transporte activo y es fundamental para la
contracción muscular.
3. Detoxificación de sustancias: En el REL hay enzimas que transforman estas
sustancias tóxicas que no son solubles en solubles, y de esta forma se eliminan
por la orina. Si la sustancia es liposoluble en vez de hidrosoluble, los hepatocitos
(son las células del hígado) aumentan su REL hasta que la eliminan. Luego el
exceso de REL es eliminado por autofagia.
4. Intervienen en el correcto plegamiento de las proteínas.
5. Detecta proteínas defectuosas, y las envía al citoplasma para que se destruyan.
• FUNCIONES DEL AG
1. Transporte y glicosilación de las proteínas procedentes del RER
2. Glicosilación de Lípidos: Para formar glucolípidos.
3. Formación de Lisosomas Primarios.
4. Formación de la Celulosa que forma la pared celular de las células eucariotas
vegetales:
Hay que tener en cuenta que la pared celular no es un orgánulo, sino que es un producto
que se excreta al exterior de la célula y que luego queda por fuera de ésta.
Solo existe en las células eucariotas vegetales y como ya sabemos la celulosa está
formada por b-D-Glucosas unidas por enlaces b( 1®4).
La pared celular tiene tres capas:
1) La laminilla media: Es la más externa de las capas. Está formada principalmente por
pectina (heteropolisacárido).
2) La pared primaria: Es la capa más interna. Está formada principalmente por
celulosa.
3) La pared secundaria: No existe siempre y cuando existe es la capa más interna.
Sólo se forma en células muy especializadas (muy maduras o muertas) y contiene
además de celulosa otras sustancias como lignina o suberina.
José Juan Canel Álvarez
21
6. FLUJOS DE ENERGÍA EN LAS CÉLULAS
Los dos orgánulos que intervienen en este flujo de energía son dos:
1. MITOCONDRIAS. 2. PLASTOS.
1. MITOCONDRIAS: Suelen ser de forma elíptica, aunque también pueden tener
forma filamentosa u oval. Su tamaño es muy pequeño (suelen ser más pequeñas que los
cloroplastos) y pueden estar en la célula en un número muy elevado en la célula (hasta
2000).
En cuanto a su estructura, es muy similar para todas las mitocondrias
independientemente de su forma y tamaño:
a. Poseen una membrana externa muy semejante a la membrana plasmática.
b. También poseen una membrana interna sin colesterol que es casi impermeable ya que
posee un fosfolípido (la cardiolipina) que la hace muy impermeable, y por lo tanto casi
todas las partículas que entran en el interior de la mitocondria lo hacen por medio de
transportadores. Además, esta membrana interna se prolonga hacia el interior formando
unos pliegues que se llaman crestas mitocondriales.
c. Espacio Intermembranal: Es el espacio que hay entre la membrana externa y la
membrana interna.
d. Matriz Mitocondrial: Es el espacio delimitado por la membrana interna. Dentro de
esta matriz mitocondrial encontramos proteínas, lípidos, ARNs, ADN doble y circular, y
ribosomas que se llaman mitorribosomas (son más pequeños que los que hay adosados
al RER y que los que hay libres en el hialoplasma). La presencia de ARN y de ADN
dentro de la mitocondria es muy importante ya que esto les permite a las mitocondrias
sintetizar alguna de sus proteínas, de manera que la mitocondria es un orgánulo
semiindependiente de la célula.
También hay que destacar que las proteínas de la membrana interna y de las crestas
mitocondriales son muy importantes en los procesos de respiración celular como
veremos cuando estudiemos el metabolismo celular.
José Juan Canel Álvarez
22
En cuanto a las funciones de las mitocondrias podemos destacar tres:
1. Oxidaciones Respiratorias.
2. Producción de Moléculas que sirven como precursores para la síntesis de
moléculas más grandes en el hialoplasma.
3. Síntesis de parte de proteínas mitocondriales.
2. LOS PLASTOS: Son orgánulos citoplasmáticos que SÓLO ESTÁN EN LOS
ORGANISMO VEGETALES. Existen varios tipos de plastos, pero debido a
su importancia nosotros en este curso sólo veremos los cloroplastos:
Son unos orgánulos muy variables en cuanto a su forma y tamaño, ya que por ejemplo
algunas células tienen sólo uno o dos cloroplastos, en cambio otras células poseen un
gran número de ellos.
Están formados por una doble membrana: La membrana externa y la membrana interna,
estando las dos membranas separadas como en la mitocondria por un espacio
Intermembranal. La membrana interna de los cloroplastos a diferencia de la de la
mitocondrias no tiene crestas y encierra un espacio central llamado estroma.
En el estroma aparecen unos sáculos que tienen pigmentos fotosintéticos y que se
llaman tilacoides. Existen dos clases de tilacoides:
1. Tilacoides del estroma o lamelas: Son de gran tamaño y sostienen a los tilacoides de
los grana.
2. Tilacoides de los grana: Son más pequeños y están apilados.
NOTA: Se llama grana al conjunto de unos tilacoides granales apilados.
Además, en el estroma están los enzimas responsables de la fase oscura de la
fotosíntesis, así como acumulaciones de almidón ( pirenoides ), y gotas de grasa (
plastoglóbulos ), copias de ADN doble y circular y ribosomas que en este caso se
llaman plastorribosomas que al igual que los de la mitocondria son más pequeños que
los ribosomas que están adheridos al RER y los ribosomas que hay libres en el
José Juan Canel Álvarez
23
hialoplasma y los cuales al igual que en las mitocondrias sintetizan una parte de las
proteínas del cloroplasto.
Las funciones más importantes de los cloroplastos son:
1. Realizan la fotosíntesis: La fase luminosa en los tilacoides y la fase oscura en
el estroma.
2. Fabrican parte de sus proteínas, siendo al igual que las mitocondrias un
orgánulo semiindependiente de la célula.
3. Intervienen en la síntesis de ácidos grasos.
4. Intervienen en la reducción de nitratos a nitritos.
Por último, decir, que tanto las mitocondrias como los cloroplastos se dividen por
bipartición, independientemente de la división de la célula.
TEORÍA ENDOSIMBIÓTICA DE MARGULIS
1. Una bacteria Arquea que utilizaba el calor como fuente de energía y que
consumía azufre, se funcionó con una bacteria nadadora y se formó la primera
célula eucariota. El núcleo se cree que se formó por fagocitosis de
Arqueobacterias.
2. Estas célula eucariotas primitivas eran anaerobias y fagocitaron células
procariotas más pequeñas que eran aerobias. Algunas de estas células
procariotas no fueron digeridas y establecieron simbiosis con las células
eucariotas primitivas y así se formaron las primeras mitocondrias y las primeras
células eucariotas aerobias. De esta incorporación proceden los animales y los
hongos.
3. Estas nuevas células aeróbicas fagocitaron bacterias fotosintéticas
(cianobacterias) y las que no fueron digeridas establecieron simbiosis con las
células aeróbicas y se formaron los cloroplastos y las primeras células eucariotas
vegetales. Así se formó el Reino Vegetal.
José Juan Canel Álvarez
24
Las pruebas más importantes a favor de esta teoría son:
1. Las mitocondrias y cloroplastos tienen ADN doble y circular al igual que las
procariotas.
2. Los ribosomas de las mitocondrias y cloroplastos son del mismo tamaño que
los de las procariotas (70S).
3. Las mitocondrias y los cloroplastos se dividen por bipartición al igual que
las procariotas.
4. Las mitocondrias y cloroplastos fabrican parte de sus proteínas.
7. FLUJOS DE INFORMACIÓN DE LA CÉLULA
Aparte del núcleo que veremos más adelante, existen unas estructuras celulares que
intervienen en este flujo y que son los ribosomas.
Los ribosomas no se ven al microscopio óptico y al electrónico son poco visibles, no
pudiendo casi ni adivinar su estructura. Están en gran número en el citoplasma y pueden
estar de tres maneras:
1. Libres en el citoplasma.
2. Adheridos al membrana del RER.
3. Dentro de las mitocondrias y de los cloroplastos.
Los ribosomas están formados por dos subunidades que sedimentan con velocidad
distinta, la cual se mide en unidades Svedberg (S), lo que indica que el ribosoma posee
una subunidad mayor y otra subunidad menor, y estas subunidades son mayor en las
células eucariotas que en las procariotas y que los de las mitocondrias y los de los
cloroplastos.
José Juan Canel Álvarez
25
Los ribosomas tienen la siguiente composición:
1. Agua. 2. ARNr.
Por otra parte, la síntesis de ARNr tiene lugar en el nucleolo, donde se origina una
molécula grande de ARN que tras su ruptura origina las dos moléculas de ARN que
forman las dos subunidades, para luego unirse con las proteínas que penetran en el
núcleo a través de los poros nucleares ya que estas proteínas en un principio están en el
hialoplasma y luego entran en el núcleo para unirse como ya dijimos al ARNr.
Por último, decir que como ya sabéis la función de los ribosomas es la síntesis de
proteínas y pueden existir multitud de ribosomas asociados a un ARNm en la síntesis de
proteínas, y a este conjunto de ribosomas recibe el nombre de Polisoma o
Polirribosomas.
José Juan Canel Álvarez
26
8. EL HIALOPLASMA
El Hialoplasma se define como el citoplasma sin orgánulos. Está constituido por
agua, sales minerales, iones y moléculas orgánicas de las que destacaremos las
proteínas. Entre las proteínas que existen en el hialoplasma podemos distinguir dos
tipos:
1. Enzimas implicados en el metabolismo celular.
2. Proteínas estructurales, las cuales forman el citoesqueleto.
Una cosa a destacar en el hialoplasma es que al poseer grandes moléculas va a sufrir
transformaciones del estado de sol a gel provocando por ejemplo el movimiento
ameboide y los fenómenos de ciclosis (es un movimiento continuo de determinados
orgánulos celulares, sobre todo de vacuolas, alrededor del núcleo celular).
EL CITOESQUELETO
Es el armazón interno de la célula ya que se une a las proteínas de la cara interna
de la membrana plasmática y es el responsable de la forma de la célula y del
movimiento celular (tanto del movimiento intracelular como del movimiento
extracelular).
El citoesqueleto está formado por tres elementos:
1. Microfilamentos de actina.
2. Microtúbulos.
3. Filamentos intermedios.
José Juan Canel Álvarez
27
1. Microfilamentos de actina:
Son imprescindibles para los movimientos celulares.
Se pueden polimerizar y despolimerizar y pueden encontrarse como monómero en
forma libre, denominada actina G, o como actina F que son dos hebras de actina G
en forma de doble hélice.
Las funciones más importantes de los filamentos de actina son:
1. Contracción muscular: En las células musculares, la actina se asocia a otra
proteína que se llama miosina, de manera que los filamentos de actina se
deslizan sobre los de miosina y esto permite la contracción muscular.
2. Citocinesis: En la citocinesis animal, se forma un anillo contráctil formado por
actina y miosina que permite la separación en dos células hijas.
3. Movimiento ameboide: Algunas organismos unicelulares, como la ameba, se
desplazan por pseudópodos o falsos pies que son prolongaciones celulares que
poseen microfilamentos de actina.
José Juan Canel Álvarez
28
2. Microtúbulos:
Los Microtúbulos son pequeños cilindros huecos, y se originan todos ellos a partir de un
Centro Organizador de Microtúbulos que es el Centrosoma o Citocentro (COMT).
Los Microtúbulos están formados por una proteína globular llamada Tubulina, en las
cuales varias unidades de Tubulina se asocian entre sí para formar un Protofilamento, y
13 Protofilamentos forman un Microtúbulo.
Las funciones más importantes de los microtúbulos son:
1. Organizan el citoesqueleto y permiten el movimiento de los orgánulos.
2. Forman el huso mitótico.
3. Forman los cilios y los flagelos.
EL CENTROSOMA, CITOCENTRO O COMT:
Se encuentra localizado cerca del núcleo y a veces está situado en una depresión de la
envoltura nuclear. Está constituido por las siguientes partes:
1. Una parte central: Los Centríolos o Diplosoma (dos centríolos = un Diplosoma).
2. Una parte brillante alrededor: La Centroesfera.
3. Una estructura filamentosa que parte de la Centroesfera: El Áster.
Los Centríolos o Diplosoma no poseen membrana y tienen
forma de barril. Son dos estructura cilíndricas situadas
perpendicularmente una a la otra, y están constituidas por
9 tripletes de microtúbulos asociados entre ellos (9+0).
Además, como ya veremos, los centríolos en la división
celular originan cada uno de ellos un centríolo nuevo, de
manera que así se obtienen dos centríolos nuevos para la
célula hija.
Las fibras del Áster y la Centroesfera durante la división
celular dan origen a los microtúbulos del huso acromático,
por lo tanto, el huso acromático aparece incluso cuando no
existen centríolos (por ejemplo, las células vegetales no
tienen centríolos, pero poseen huso mitótico el cual
forman a partir de una estructura que se llama estructura
difusa).
José Juan Canel Álvarez
29
Las funciones más importantes de los centríolos son:
1. Formar el huso mitótico en células animales.
2. Intervienen en la formación de los cilios y flagelos.
3. Se duplican en el período S de la fase de interfase, para que cada célula hija
tenga dos centríolos.
4. La posición de los centríolos determina la posición del núcleo celular
3. Filamentos intermedios:
Se llaman así porque su diámetro es intermedio entre los microfilamentos de actina
y los microtúbulos.
Están formados por proteínas fibrosas, forman redes alrededor del núcleo y se van
extendiendo hacia la periferia.
Los filamentos intermedios se pueden agrupar en tres clases:
1. Filamentos de queratina: también se llaman tonofilamentos. Confieren una
elevada resistencia mecánica.
2. Neurofilamentos: Están dentro de las neuronas.
3. Filamentos de vimentina (en células como fibroblastos), desmina (en células
musculares) y GFAP (en astrocitos).
Los filamentos intermedios son los más resistentes de los tres y realizan funciones
estructurales, evitando la rotura de las membranas de las células que se encuentran
sometidas a tensiones mecánicas. Además, junto al resto del citoesqueleto ayudan a
mantener la forma celular.
CILIOS Y FLAGELOS
Son prolongaciones del citoplasma, se forman a partir de los centriolos y cuya función
es permitir el movimiento de la célula.
Los cilios son muy cortos y numeroso, en cambio los flagelos son largos y pocos,
aunque los dos tienen la misma estructura:
José Juan Canel Álvarez
30
1. Tallo o axonema:
Está formados por 9 pares de microtúbulos asociados entre sí por puentes entre
las proteínas de los microtúbulos más dos centrales. Esto se conoce como
estructura 9+2
• Los dos microtúbulos centrales son completos, es decir cada microtúbulo tiene
13 protofilamentos.
• En cambio, los periféricos, el microtúbulo A es completo, pero el B sólo tiene 10
protofilamentos (comparte con el microtúbulo A tres protofilamentos).
El microtúbulo A tiene dos prolongaciones formadas por una proteína que se llama
dineína que es una ATPasa que permite el deslizamiento de unas parejas de
microtúbulos sobre otros y las parejas de microtúbulos adyacentes están unidas por una
proteína que se llama nexina.
José Juan Canel Álvarez
31
2. Zona de transición: Desaparece el par de microtúbulos centrales.
3. Corpúsculo basal: Tiene la misma estructura que el centríolo: 9+0.
El microtúbulo A es completo, mientras que B y C son incompletos y tiene 10
protofilamentos.
Las células vegetales como no tiene centríolos, los cilios y flagelos los
desarrollan a partir del corpúsculo basal.
4. Raíces ciliares: Son microfilamentos que salen del extremo inferior del
corpúsculo basal y cuya función es la coordinación del movimiento de los cilios.
9. EL NÚCLEO
Es el orgánulo celular que controla y gobierna todas las funciones de la célula y donde
se encierra la mayor parte del material hereditario (el ADN).
SÓLO EXISTE EN LAS CÉLULAS EUCARIOTAS y se puede presentar con dos
aspectos muy distintos:
a. Núcleo en Interfase. b. Núcleo en división.
En este tema vamos a estudiar el núcleo en interfase (la interfase es el período que hay
entre dos divisiones celulares seguidas), y también se le llama núcleo en reposo, lo que
no quiere decir que no posea actividad, sino al contrario, ya que la interfase es una etapa
de gran actividad, pero se llama así porque no se está dividiendo.
• Aspecto: Suele tener forma de esfera y suele estar separado del citoplasma por una
membrana nuclear (recordar que esta envoltura nuclear se continuaba con el RER).
El contenido del núcleo se va más o menos homogéneo salvo por la presencia de
unas pequeñas estructuras esféricas llamadas nucleolos.
• Número: Generalmente las células suelen presentar un solo núcleo, pero podemos
poner el ejemplo del paramecio, el cual posee dos núcleos, uno mayor llamado
macronúcleo, y otro menor llamado micronúcleo. Otro ejemplo puede se los
Osteoclastos (son células óseas), las cuales poseen numerosos núcleos.
José Juan Canel Álvarez
32
• Forma: Puede ser esférico, elíptico, irregular, etc. Por ejemplo, los glóbulos blancos
tienen un núcleo lobulado muy irregular.
• Tamaño: Es bastante constante en una estirpe celular, Es muy voluminosos en las
células que todavía no están diferenciadas o en las que están muy activas. Si el
núcleo sufre aumento de volumen es un indicio de que la célula está a punto de
entrara en división.
• Ultraestructura: Al microscopio electrónico podemos distinguir los siguientes
elementos:
1. Envoltura o Membrana nuclear:
Es doble y entre la membrana externa y la interna hay un espacio que se llama
espacio perinuclear. quedando entre ellas un espacio llamado espacio perinuclear. La
envoltura nuclear no es continua, sino que tiene un gran número de poros.
Estos complejos de poros están formados por 8 gránulos proteicos, de tal manera que
estos complejos permiten el paso de grandes moléculas e impiden que se produzcan
diferencias osmóticas entre el núcleo y el citoplasma.
En el interior del núcleo y adosada a la membrana interna se encuentra unas proteínas
que se llaman Lámina Densa o nuclear, siendo su función principal la de formar el
núcleo, de manera que en las mitosis estas láminas densas se fosforilan y se separan por
lo que el núcleo se desorganiza. Una vez concluida la mitosis, estas láminas se
desfosforilan y se vuelven a unir, por lo que el núcleo se vuelve a organizar.
2. Nucleoplasma: Es el jugo nuclear, y en cuanto a su composición es semejante al
hialoplasma, pero no posee microtúbulos, ni microfilamentos. Está formado por agua,
proteínas, ARN e iones. En el nucleoplasma se produce la síntesis del ARN
(transcripción), además inmersos en el nucleolo encontramos el nucleolo y la cromatina.
3. Cromatina: Recibe este nombre ya que se tiñe con colorantes básicos. Está
constituida básicamente por ADN, proteínas y algo de ARN.
Estas fibras de cromatina como ya sabemos tienen distintos niveles de organización
(nucleosoma, collar de perlas..., cromosoma) de los cuales ya hemos hablado en el tema
de Ácidos Nucleicos, y que permiten empaquetar grandes cantidades de ADN en un
espacio muy reducido como es el núcleo celular.
Las fibras de cromatina constituyen los cromosomas interfásicos, los cuales se
encuentran tan extendidos y enmarañados que resulta imposible distinguirlos. Lugo,
durante la fase S, como ya veremos en el tema de mitosis, el ADN se duplica y cada
cromosoma origina una copia idéntica de sí mismo, por lo que a partir de este momento
cada cromosoma consta de dos subunidades idénticas llamadas cromátidas.
En el núcleo existen dos clases de cromatina:
1. Eucromatina: Son partes de la cromatina que están poco condensadas y por lo tanto
en estas zonas se puede realizar la transcripción. Por lo tanto, la eucromatina junto con
los nucleolos son zonas donde se transcriben los genes.
José Juan Canel Álvarez
33
2. Heterocromatina: Es la parte de la cromatina que está muy empaquetada, de manera
que esta parte de ADN no se transcribe. De esta heterocromatina existen dos clases:
2.1. Heterocromatina Constitutiva: El ADN está siempre condensado y los genes nunca
se transcriben. Se desconoce su función, aunque se cree que está relacionada con la
meiosis en el apareamiento de los cromosomas homólogos.
2.2. Heterocromatina Facultativa: Son zonas de la cromatina que en una clase de células
puede estar inactivada y otra clase de células no.
4. Nucleolo: Es una estructura esférica, que se destaca del resto del contenido nuclear
por ser más brillante. Aparece con frecuencia asociado a zonas de cromatina que son los
organizadores nucleolares. El nucleolo no es constante en número y depende del estado
funcional de la célula, ya que por ejemplo cuando la célula se va a dividir desaparece.
En cuanto a su estructura no presenta membrana de separación con el núcleo, y está
básicamente constituido por ARN, ya que, como ya sabemos, es el nucleolo donde se va
a dar la síntesis de los ARN ribosomales.
José Juan Canel Álvarez
34
10. DIFERENCIAS ENTRE UNA CÉLULA ANIMAL Y UNA CÉLULA
VEGETAL
CÉLULA ANIMAL CÉLULA VEGETAL
TAMAÑO MEDIO MENOR MAYOR
PARED CELULAR NO SÍ, FORMADA POR
CELULOSA
PLASTOS NO SÍ
CENTRÍOLOS SÍ NO, SALVO EXCEPCIONES
MUY RARAS
VACUOLAS MÁS PEQUEÑAS Y MÁS EN MENOR NÚMERO Y
NUMEROSAS MÁS GRANDES
CILIOS Y FLAGELOS FRECUENTES NO SALVO EXCEPCIONES
RESERVA GLUCÓGENO ALMIDÓN.
HIDROCARBONADA
POSICIÓN DEL NÚCLEO CENTRAL EXCÉNTRICO
INCLUSIONES GLUCÓGENO, LÍPIDOS LÁTEX, LÍPIDOS...
NUTRICIÓN HETERÓTROFA AUTÓTROFA
FOTOSINTÉTICA, SALVO
EXCEPCIONES (RAÍZ ...)
MOVIMIENTO POR CILIOS, FLAGELOS, NO, SALVO EXCEPCIONES
PSEUDÓPODOS
MITOSIS ASTRAL. LOS ANASTRAL. LOS
MICROTÚBULOS DEL HUSO MICROTÚBULOS DEL HUSO
SE ORGANIZAN A PARTIR SE ORGANIZAN A PARTIR
DE LOS CENTRÍOLOS DE UNA ZONA NO
DIFERENCIADA DEL
CITOPLASMA LLAMADA
ESTRUCTURA DIFUSA
DIVISIÓN DEL POR ESTRANGULAMIENTO POR CRECIMIENTO DE UN
CITOPLASMA TABIQUE LLAMADO
(=CITOCINESIS) FRAGMOPLASTO
José Juan Canel Álvarez
35
CLASES DE MICROSCOPIOS
A) MICROSCOPIO ÓPTICO O FOTÓNICO
A.1) FUNDAMENTO
Funciona de la siguiente manera: Una fuente luminosa envía rayos de luz a una primera
lente, llamada condensador, que concentra los rayos de luz sobre el objeto a observar.
Estos rayos atraviesan el objeto y una lente denominada objetivo da una imagen
aumentada de éste. Una segunda lente, el ocular, vuelve a aumentar la imagen dada por
el objetivo. Esta última imagen es la que será recibida por el observador.
• El poder de resolución es la distancia mínima a la que tienen que estar dos
puntos de la muestra para que puedan verse como distintos.
• Para calcular los aumentos a los que observamos se multiplica el aumento
del ocular por el del objetivo.
A.2) PREPARACIÓN DEL MATERIAL
En el microscopio óptico la luz atraviesa el objeto a observar. Si éste es muy grueso, la
luz no lo atravesará y el objeto aparecerá demasiado oscuro. Si el objeto es demasiado
delgado o muy transparente, no se observarán sus estructuras. En cualquier caso,
deberemos realizar una preparación.
En general, una preparación requiere las siguientes etapas
1- CORTE. Los objetos son cortados mediante aparatos denominados microtomos.
El tejido destinado al corte debe congelarse o incluirse en parafina para darle una
mayor consistencia y que se pueda cortar con facilidad.
José Juan Canel Álvarez
36
2- FIJACIÓN. Su fin es matar a las células con la menor alteración de las estructuras
posible. Como fijadores se emplean determinadas sustancias químicas (por ejemplo:
formaldehído y tetróxido de osmio).
3- DESHIDRATACIÓN. La extracción del agua del interior de las células permitirá
también una mejor conservación y la penetración de los colorantes. Para deshidratar
el material a observar se le sumerge en alcoholes de cada vez mayor graduación que
por dilución irán extrayendo el agua.
4- TINCIÓN. Es la coloración de las células o de partes de éstas para que resalten y
posibilitar así su observación. Algunos colorantes son selectivos pues tiñen partes
concretas de la célula.
Existen dos clases de colorantes:
a) Los colorantes vitales. Que tiñen las estructuras celulares pero sin matar a
las células (por ejemplo: el verde jano, el rojo neutro, el azul tripán, el azul de
metileno).
b) Los colorantes no vitales. Que matan a las células (eosina, hematoxilina).
5- MONTAJE. Una vez realizadas las anteriores operaciones el material se coloca
entre un porta-objetos y un cubre-objetos. Para un montaje no definitivo, se coloca
entre "porta" y "cubre" una gota de glicerina. Este tipo de preparaciones tiene una
duración limitada y sólo sirven para la observación momentánea o a lo sumo de unos
días. Si se desea una mayor duración debe realizarse el montaje en gelatina-glicerina
o en euparal.
B) EL MICROSCOPIO ELECTRÓNICO
Existen dos clases de microscopios electrónicos:
B.1) Microscopio electrónico de trasmisión (MET).
B.2) Microscopio electrónico de barrido (MEB).
B.1) EL MICROSCOPIO ELECTRÓNICO DE TRANSMISIÓN (MET)
FUNDAMENTO No se puede mostrar la imagen.
En esencia su funcionamiento es similar al del
microscopio óptico. Un cátodo emite un haz de
electrones que son acelerados por la aplicación de una
diferencia de potencial entre el cátodo y el ánodo. El
flujo de electrones es concentrado sobre el objeto por una
primera lente magnética que hace las veces de
condensador. Los electrones atraviesan la muestra. Una
segunda lente magnética, el objetivo, da una imagen
aumentada del objeto. Una tercera lente, el ocular,
José Juan Canel Álvarez
37
aumenta de nuevo la imagen dada por la anterior. La imagen final es proyectada sobre
una pantalla o fotografiada.
B.1.2) PREPARACIÓN del MATERIAL
Los electrones necesitan desplazarse en el vacío, esta es la razón por la que no es
posible la observación de células vivas al microscopio electrónico.
1º) FIJACIÓN. Las células son fijadas mediante fijadores no coagulantes. Los más
fijadores más corrientes son corrientes son el tetróxido de osmio (OsO4), el
formaldehído (HCHO) y el permanganato potásico (MnO4K).
2º) DESHIDRATACIÓN e INCLUSIÓN. La pieza es deshidratada e infiltrada
mediante una resina o plástico para darle una mayor consistencia y facilitar su corte.
3º) CORTE. Los cortes se realizan mediante ultramicrotomos de cuchilla de vidrio
o de diamante.
B.2) MICROSCOPIO ELECTRÓNICO DE BARRIDO (MEB)
Este tipo de microscopio permite obtener imágenes tridimensionales del objeto a
estudiar.
José Juan Canel Álvarez
38
José Juan Canel Álvarez
39
José Juan Canel Álvarez
40
7.
8.
9.
José Juan Canel Álvarez
41
10.
11.
12.
José Juan Canel Álvarez
42
13.
14.
José Juan Canel Álvarez
También podría gustarte
- Semana 4 - La CelulaDocumento67 páginasSemana 4 - La CelulaEliane TrinidadAún no hay calificaciones
- Taller No 123 B.molecularDocumento29 páginasTaller No 123 B.molecularDavid EstebanAún no hay calificaciones
- Clase 3 Organizacià N CelularDocumento58 páginasClase 3 Organizacià N CelularTairaAún no hay calificaciones
- Ciencias Naturales PracticaDocumento8 páginasCiencias Naturales Practicajosuedejesus321Aún no hay calificaciones
- BIO Sem3 Citología IDocumento7 páginasBIO Sem3 Citología IJennifer PerezAún no hay calificaciones
- 3-La CelulaDocumento6 páginas3-La CelulaAlejandra Masgoret CuellarAún no hay calificaciones
- Tema 1.-Citologia Y Organización Funcional de La Vida: MSC - Dra.Vania Salazar Rendon - BiologíaDocumento8 páginasTema 1.-Citologia Y Organización Funcional de La Vida: MSC - Dra.Vania Salazar Rendon - BiologíaBohemia BoAún no hay calificaciones
- Semana 6 - Biología Celular I - Anual Sm.Documento82 páginasSemana 6 - Biología Celular I - Anual Sm.kamyla quispeAún no hay calificaciones
- Clase 04 Estructura Celular Parte 1Documento26 páginasClase 04 Estructura Celular Parte 1Bigtaesty ThinkAún no hay calificaciones
- Histologia Practica 3Documento7 páginasHistologia Practica 3Iliana ZapotAún no hay calificaciones
- Revista Sobre La CelulaDocumento9 páginasRevista Sobre La CelulaYoselin HdzAún no hay calificaciones
- Csa Biologia 10 CelulasDocumento7 páginasCsa Biologia 10 CelulasGianna LezcanoAún no hay calificaciones
- U2 Act 2 Estructura Física de La Celula.Documento19 páginasU2 Act 2 Estructura Física de La Celula.Manuel RodriguezAún no hay calificaciones
- La Celula para Quinto GradoDocumento3 páginasLa Celula para Quinto GradoAndrea Pita RequenaAún no hay calificaciones
- Celular VerDocumento26 páginasCelular VerJennyfer QuirozAún no hay calificaciones
- BIO - Sem3 - Citología IDocumento8 páginasBIO - Sem3 - Citología Igianella thirza salazar huamanAún no hay calificaciones
- Tema 6 La Célula: Bloque II.-La Célula Viva. Morfología, Estructura y Fisiología CelularDocumento23 páginasTema 6 La Célula: Bloque II.-La Célula Viva. Morfología, Estructura y Fisiología CelularVanessa Lorenzo PérezAún no hay calificaciones
- Clase 2 - BiologíaDocumento45 páginasClase 2 - BiologíaSandraSalazarAún no hay calificaciones
- Apunte de Biologia Fundamentos de La BioDocumento31 páginasApunte de Biologia Fundamentos de La BioRon RuizAún no hay calificaciones
- SESIONDocumento6 páginasSESIONLucio Rojas VitorAún no hay calificaciones
- BioquimicaDocumento175 páginasBioquimicaSANTIAGO ANDRÉS MASMUTA PUPIALESAún no hay calificaciones
- Unidad 3 Fisiologia Celular (Nuevo)Documento26 páginasUnidad 3 Fisiologia Celular (Nuevo)lme7692Aún no hay calificaciones
- La Célula, Biologia.Documento3 páginasLa Célula, Biologia.Astrith Andrea Tellez NajarroAún no hay calificaciones
- Taller de La CélulaDocumento4 páginasTaller de La CélulajohncastaveAún no hay calificaciones
- Bioquímica. La Organización CelularDocumento11 páginasBioquímica. La Organización CelularConsuelo Zurita RicoAún no hay calificaciones
- Células Ecuariotas y ProcariotasDocumento7 páginasCélulas Ecuariotas y ProcariotasJhoel ChAún no hay calificaciones
- La CélulaDocumento14 páginasLa CélulaRodrigo Fernandez DominguezAún no hay calificaciones
- TEMA 8 - Organizacion y Estructura CelularDocumento7 páginasTEMA 8 - Organizacion y Estructura CelularproyventAún no hay calificaciones
- Clase 1 Biol 130 012020 Introduccion A Las Celulas Parte IDocumento51 páginasClase 1 Biol 130 012020 Introduccion A Las Celulas Parte Icamila palominoAún no hay calificaciones
- Guia 8 Biologia 10Documento4 páginasGuia 8 Biologia 10fernando mur saenzAún no hay calificaciones
- Captura de Pantalla 2022-12-17 A La(s) 9.10.29 P.M.Documento10 páginasCaptura de Pantalla 2022-12-17 A La(s) 9.10.29 P.M.Alondra MaldonadoAún no hay calificaciones
- GUIA 1° Medio BIOLOGIA Celula 2010Documento5 páginasGUIA 1° Medio BIOLOGIA Celula 2010gloritaalgarroboAún no hay calificaciones
- U2 Act 2 Estructura Fisica de La CelulaDocumento15 páginasU2 Act 2 Estructura Fisica de La CelulaKAROL GUTIERREZ MADRIGALAún no hay calificaciones
- TALLER DE RECUPERACION DE CIENCIAS NATURALES CURSO 603 DE Vladimir OliveraDocumento4 páginasTALLER DE RECUPERACION DE CIENCIAS NATURALES CURSO 603 DE Vladimir Oliverawilliam martinezAún no hay calificaciones
- Fisiopatologia GeneralDocumento6 páginasFisiopatologia GeneralMeritxell0% (1)
- La Célula: Q.F Miriam A. Enriquez SaireDocumento25 páginasLa Célula: Q.F Miriam A. Enriquez SaireMIRIAM AYDE ENRIQUEZ SAIREAún no hay calificaciones
- T 6 La CelulaDocumento24 páginasT 6 La CelulaRaul Ibanez RodriguezAún no hay calificaciones
- Resumen Final de Biología Celulas (Editado)Documento12 páginasResumen Final de Biología Celulas (Editado)Galilea Aráoz DíazAún no hay calificaciones
- ACTIVIDADES DE REPASO DEL TEMA 1. Parte 1Documento5 páginasACTIVIDADES DE REPASO DEL TEMA 1. Parte 1Javier DelgadoAún no hay calificaciones
- Anatomia y Fisiología CelularDocumento56 páginasAnatomia y Fisiología CelularKatherin VillanuevaAún no hay calificaciones
- Repaso Primer Parcial BioquimicaDocumento29 páginasRepaso Primer Parcial BioquimicaKary AyonAún no hay calificaciones
- BioquímicaDocumento6 páginasBioquímicamaria chaconAún no hay calificaciones
- Hoja de Trabajo 2 OrganelosDocumento4 páginasHoja de Trabajo 2 OrganelosStu StuAún no hay calificaciones
- Tipos de CélulasDocumento7 páginasTipos de CélulasKrlitos AmadorAún no hay calificaciones
- Guía de Laboratorio - Biología Celular - Profesora Deinorah - 2022-3Documento9 páginasGuía de Laboratorio - Biología Celular - Profesora Deinorah - 2022-3Jessy MojicaAún no hay calificaciones
- La CelulaDocumento15 páginasLa Celulaesme gonzalezAún no hay calificaciones
- Cartilla Celula EucariotaDocumento6 páginasCartilla Celula EucariotaMâytë TämbøAún no hay calificaciones
- Tema 1 Or... 1Documento17 páginasTema 1 Or... 1MONICA SALMERON MONTEROAún no hay calificaciones
- Anatomía CelularDocumento35 páginasAnatomía CelularLrNCrZAún no hay calificaciones
- Qué Es La CélulaDocumento5 páginasQué Es La Célulajhoseppacco4Aún no hay calificaciones
- El Estudio de La CelulaDocumento20 páginasEl Estudio de La CelulaLuis Miguel Leonardo FabianAún no hay calificaciones
- La CélulaDocumento36 páginasLa CélulaHernan CarrascoAún no hay calificaciones
- BiologíaDocumento11 páginasBiologíaMartín BarottoAún no hay calificaciones
- La Celula.Documento25 páginasLa Celula.jose antonio soria paredesAún no hay calificaciones
- Célula - HistologíaDocumento108 páginasCélula - Histologíakaren pennisiAún no hay calificaciones
- Clase 3 Teoria La CelulaDocumento81 páginasClase 3 Teoria La CelulaJhestynAnyeroToribioCarlosAún no hay calificaciones
- Tema 1 Organizacion General de Las CelulasDocumento39 páginasTema 1 Organizacion General de Las CelulasSarah Jessica Quiroz OlivaresAún no hay calificaciones
- La Célula ExpoDocumento54 páginasLa Célula ExpoMarco Antonio Rivera PAún no hay calificaciones
- Preguntas AnatomiaDocumento54 páginasPreguntas Anatomiamirna victoria reynoso lopezAún no hay calificaciones
- Borrador Examen FisicoDocumento18 páginasBorrador Examen FisicoAndres Santiago Ramirez SalamancaAún no hay calificaciones
- Recopilación Preguntas Certamen 1 (Lesión-Inflamación-Inmune-Cáncer) - 1Documento18 páginasRecopilación Preguntas Certamen 1 (Lesión-Inflamación-Inmune-Cáncer) - 1bruno martinezAún no hay calificaciones
- CARVER - Quien Quiera Que Hubiera Dormido 1Documento9 páginasCARVER - Quien Quiera Que Hubiera Dormido 1Pablo Eugenio FernandezAún no hay calificaciones
- El Cerebro y La ConductaDocumento6 páginasEl Cerebro y La ConductaJohanny TaverasAún no hay calificaciones
- Relaciones Con La Odontología (Trabajo Final de Fisiologia)Documento3 páginasRelaciones Con La Odontología (Trabajo Final de Fisiologia)BisMark MendozaAún no hay calificaciones
- Guía de Estudio Tercer Parcial FisiologíaDocumento7 páginasGuía de Estudio Tercer Parcial FisiologíadiegojestradatrejoAún no hay calificaciones
- Clase 9 04-10Documento2 páginasClase 9 04-10Lismaryora GodoyAún no hay calificaciones
- APUNTE Mecanica Corporal y Movilización-1Documento27 páginasAPUNTE Mecanica Corporal y Movilización-1eliana rodriguezAún no hay calificaciones
- Control de Temperatura ECAP VIERNES 20-11-2020-1Documento10 páginasControl de Temperatura ECAP VIERNES 20-11-2020-1Ernesto gomez aguirreAún no hay calificaciones
- Sesion 4 Sistema Digestivo 1Documento54 páginasSesion 4 Sistema Digestivo 1marlon llanos basurtoAún no hay calificaciones
- Colgajos CutáneosDocumento11 páginasColgajos CutáneosKarla Rojas NoeAún no hay calificaciones
- Mecanica Ondulatorio.Documento5 páginasMecanica Ondulatorio.Fernanda Lopez LaureanAún no hay calificaciones
- Biomecanica de Los Ligamentos.Documento11 páginasBiomecanica de Los Ligamentos.JHENIFER BIANCA VILLARROEL ILLATARCOAún no hay calificaciones
- Fisicoquimica - La UbreDocumento9 páginasFisicoquimica - La UbreCarolina RodriguesAún no hay calificaciones
- La Descripcion de Todas Las Emociones (Aversión, Tristeza, Etc) .Documento3 páginasLa Descripcion de Todas Las Emociones (Aversión, Tristeza, Etc) .Jeremy RenojoAún no hay calificaciones
- Introduccion A Sistema Inmune e Inmunidad InnataDocumento60 páginasIntroduccion A Sistema Inmune e Inmunidad InnataYerelin Rebeca Valverde Grijalva (Yere)Aún no hay calificaciones
- Plan de Actividades Semana 24 - C y T (14!09!2020)Documento5 páginasPlan de Actividades Semana 24 - C y T (14!09!2020)Fidel FloresAún no hay calificaciones
- El Abdomen - ActualizadoDocumento9 páginasEl Abdomen - ActualizadoRicardo Agudelo Giraldo50% (2)
- Actividad 1Documento8 páginasActividad 1edison renan quispe ramosAún no hay calificaciones
- Bioquimica Folleto CarreraDocumento4 páginasBioquimica Folleto CarreraCristinaKnuckeyZúñigaAún no hay calificaciones
- Clase 6. NF I. Sistemas Sensitivos - Sistema Sensitivo SomaticoDocumento14 páginasClase 6. NF I. Sistemas Sensitivos - Sistema Sensitivo SomaticoLissett CarvajalAún no hay calificaciones
- Sistema Digestivo BovinoDocumento8 páginasSistema Digestivo BovinoAllison BejaranoAún no hay calificaciones
- 1 BiologíaDocumento17 páginas1 Biologíadayanna wingAún no hay calificaciones
- Elongación de CuádricepsDocumento6 páginasElongación de Cuádricepsjacquelineandre16Aún no hay calificaciones
- Clasificacion de Las HormonasDocumento42 páginasClasificacion de Las HormonasGianella Fajardo100% (3)
- 9-Tecnicas complementarias-MTCh-VENTOSASDocumento8 páginas9-Tecnicas complementarias-MTCh-VENTOSASvr46hdsAún no hay calificaciones
- Cornea Mapa MentalDocumento1 páginaCornea Mapa MentalDulce Maria Betancourt100% (1)
- Boletín 0 SM Intensivo 2022-IDocumento59 páginasBoletín 0 SM Intensivo 2022-IJOSUE EDMUNDO BENDEZU AGUILARAún no hay calificaciones
- Habitos OralesDocumento55 páginasHabitos OralesMaria Lucrecia Quino Montes100% (1)