También podría gustarte
- Resistencia A La InsulinaDocumento25 páginasResistencia A La InsulinaLUISA FERNANDA AGUIILAR ROMANAún no hay calificaciones
- Membrana CelularDocumento4 páginasMembrana CelularJeffrey Rodriguez100% (4)
- MecanismosMolecularesRIDocumento15 páginasMecanismosMolecularesRIAlejandro Hernandez GuzmanAún no hay calificaciones
- Anemias Secundarias A Padecimientos No HematlogicosDocumento51 páginasAnemias Secundarias A Padecimientos No HematlogicosDiana Isabel AguilarAún no hay calificaciones
- Nutrigenética y Nutrigenómica: La Revolución Sanitaria Del Nuevo Milenio. Implicancias Clínicas en Síndrome Metabólico y Diabetes Tipo 2Documento8 páginasNutrigenética y Nutrigenómica: La Revolución Sanitaria Del Nuevo Milenio. Implicancias Clínicas en Síndrome Metabólico y Diabetes Tipo 2Gerson Andres RojasAún no hay calificaciones
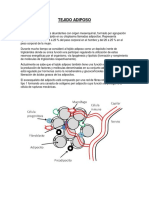
- Tejido AdiposoDocumento7 páginasTejido AdiposoJulinho M LeOnAún no hay calificaciones
- ¿Puede el cannabis ayudarte a perder peso?De Everand¿Puede el cannabis ayudarte a perder peso?Aún no hay calificaciones
- Octeto Ominoso de La DiabetesDocumento10 páginasOcteto Ominoso de La DiabetesNicolas Torres100% (4)
- Muscle Lipid Seminario2.en - EsssDocumento11 páginasMuscle Lipid Seminario2.en - EsssDaniel Andrey Franco BejaranoAún no hay calificaciones
- Fung, L., Et Al. (2015) - Resistencia A La Insulina en La MujerDocumento12 páginasFung, L., Et Al. (2015) - Resistencia A La Insulina en La MujerTamara BravoAún no hay calificaciones
- Macrófagos, Inflamación, Tejido Adiposo, Obesidad y Resistencia A La InsulinaDocumento8 páginasMacrófagos, Inflamación, Tejido Adiposo, Obesidad y Resistencia A La Insulinaphouses101100% (1)
- Adipocitos, Obesidad Visceral, Inflamación y EnfermedadDocumento7 páginasAdipocitos, Obesidad Visceral, Inflamación y EnfermedadAna Gariza SolanoAún no hay calificaciones
- Adiponectina-El Tejido Adiposo Mas Alla de La Reserva Inerte de EnergiaDocumento7 páginasAdiponectina-El Tejido Adiposo Mas Alla de La Reserva Inerte de EnergiamediquitaAún no hay calificaciones
- Nutrigenética y NutrigenómicaDocumento13 páginasNutrigenética y NutrigenómicaFer PauthAún no hay calificaciones
- Pia Articulos de Rev - TSDN - PDFDocumento22 páginasPia Articulos de Rev - TSDN - PDFOla Conde VIIAún no hay calificaciones
- Síndrome X en rumiantesDocumento15 páginasSíndrome X en rumiantesMARCOS EDUARDO MARQUEZ MARTINEZAún no hay calificaciones
- García-Torres Et Al, 2011. ESPDocumento21 páginasGarcía-Torres Et Al, 2011. ESPXochitemiqui YoacihualtAún no hay calificaciones
- Inflamación obesidad metabolismoDocumento6 páginasInflamación obesidad metabolismoAna Gabriela PuiconAún no hay calificaciones
- Síndrome Metabólico - Héctor MoralesDocumento18 páginasSíndrome Metabólico - Héctor MoralesMauricio MoralesAún no hay calificaciones
- Adipocitoquinas:mediadoresclaveeninflamaciónDocumento19 páginasAdipocitoquinas:mediadoresclaveeninflamaciónLidia RamirezAún no hay calificaciones
- Expo - Sem7 - Histologia SeminarioDocumento14 páginasExpo - Sem7 - Histologia SeminarioOsver Jahir Guevara DávilaAún no hay calificaciones
- AdipocitocinasDocumento8 páginasAdipocitocinasCuenta GoogleAún no hay calificaciones
- Informe 2 Histología SeminarioDocumento8 páginasInforme 2 Histología SeminarioEmery Sofía C. D. BustamanteAún no hay calificaciones
- Ane223e UnlockedDocumento9 páginasAne223e UnlockedLuismi DocAún no hay calificaciones
- Fisiologia Del Tejido Adiposo LipidosDocumento10 páginasFisiologia Del Tejido Adiposo LipidosStefany TorresAún no hay calificaciones
- Presentación Resistencia InsulinaDocumento16 páginasPresentación Resistencia InsulinaNerea Trillo PérezAún no hay calificaciones
- Síndrome MetabólicoDocumento33 páginasSíndrome MetabólicoHernan RoseroAún no hay calificaciones
- 3641-Texto Del Artículo-13678-1-10-20210709Documento7 páginas3641-Texto Del Artículo-13678-1-10-20210709Roger Caruajulca Cirujano GeneralAún no hay calificaciones
- Temas Del Módulo Endócrino - MetabólicoDocumento50 páginasTemas Del Módulo Endócrino - MetabólicoAtenea EncuadernacionesAún no hay calificaciones
- Entrenamiento Intervlico de Alta Intensidad HIITDocumento8 páginasEntrenamiento Intervlico de Alta Intensidad HIITMayeAún no hay calificaciones
- Sindrome MetabolicoDocumento27 páginasSindrome MetabolicoCh RoAún no hay calificaciones
- er033eDocumento7 páginaser033evsifontes91Aún no hay calificaciones
- Resistencia A La InsulinaDocumento7 páginasResistencia A La InsulinaAdriana AdameAún no hay calificaciones
- Moléculas Que Influyen en La ObesidadDocumento4 páginasMoléculas Que Influyen en La ObesidadMirella MendozaAún no hay calificaciones
- InsulinaDocumento6 páginasInsulinaPaxZAún no hay calificaciones
- El Papel de Los Acidos Grasos Libres en La ResisteDocumento13 páginasEl Papel de Los Acidos Grasos Libres en La ResisteF CPAún no hay calificaciones
- Genes asociados a obesidadDocumento18 páginasGenes asociados a obesidadclaudiaAún no hay calificaciones
- 6-Resistencia A La Insulina.Documento46 páginas6-Resistencia A La Insulina.pablonicofernandezAún no hay calificaciones
- Nutrigenética y nutrigenómica: la revolución sanitaria del nuevo milenioDocumento8 páginasNutrigenética y nutrigenómica: la revolución sanitaria del nuevo milenioPAULA SERVINAún no hay calificaciones
- Sindrome MetabolicoDocumento50 páginasSindrome MetabolicoL Taty RuizAún no hay calificaciones
- Síndrome MetabolicoDocumento37 páginasSíndrome MetabolicoITZAMA YAMILET HERNANDEZ REYNOSOAún no hay calificaciones
- Actualizacion en ObesidadDocumento8 páginasActualizacion en ObesidadadelaidaanguenvemengueAún no hay calificaciones
- GMM_2017 Resistencia InsulinaDocumento15 páginasGMM_2017 Resistencia InsulinaLucero GutierrezAún no hay calificaciones
- Obesidad y tejido adiposo blancoDocumento12 páginasObesidad y tejido adiposo blancoCarla NuñezAún no hay calificaciones
- INFORME S2 - Grupo 04 - Histo SeminarioDocumento10 páginasINFORME S2 - Grupo 04 - Histo SeminarioStephany CastroAún no hay calificaciones
- Adiponectina y Sus Efectos Pleiotrópicos en El Sistema Cardiovascular. RevisiónDocumento8 páginasAdiponectina y Sus Efectos Pleiotrópicos en El Sistema Cardiovascular. Revisión21121c0514Aún no hay calificaciones
- Proteinas DesacoplantesDocumento7 páginasProteinas DesacoplantesNicolás Sanhueza PetitAún no hay calificaciones
- Tema 8 Resistencia A La Insulina Equipo 4Documento25 páginasTema 8 Resistencia A La Insulina Equipo 4LUISA FERNANDA AGUIILAR ROMANAún no hay calificaciones
- SX MetabolicoDocumento29 páginasSX MetabolicoJeremy ChonilloAún no hay calificaciones
- Sindrome Metabolico y DiabetesDocumento6 páginasSindrome Metabolico y DiabetesEnrique Hernandez Martinez100% (1)
- Las Adipocinas y Su Importancia en El EnvejecimientoDocumento5 páginasLas Adipocinas y Su Importancia en El EnvejecimientoGian Carlos Chafloque GuzmanAún no hay calificaciones
- Efectos Pleiotrópicos de Los Inhibidores Del SGLT-2 en La Salud Cardiometabólica de Los Pacientes Con Diabetes Mellitus Tipo 2Documento16 páginasEfectos Pleiotrópicos de Los Inhibidores Del SGLT-2 en La Salud Cardiometabólica de Los Pacientes Con Diabetes Mellitus Tipo 2Luis Carlos OlivarAún no hay calificaciones
- Tejido Adiposo Como Organo Endocrino PDFDocumento9 páginasTejido Adiposo Como Organo Endocrino PDFManuel GonzalezAún no hay calificaciones
- Síndrome MetabólicoDocumento47 páginasSíndrome MetabólicoDanae HernandezAún no hay calificaciones
- OBESIDADDocumento25 páginasOBESIDADRAFAEL TORTOSAAún no hay calificaciones
- Diabetes tipo 2: FisiopatologíaDocumento9 páginasDiabetes tipo 2: FisiopatologíaWilbert Quispe MaytaAún no hay calificaciones
- 2.3.1 Obesidad abordaje clìnico N pediatrìcaDocumento19 páginas2.3.1 Obesidad abordaje clìnico N pediatrìcagfwjmh8nk6Aún no hay calificaciones
- Avance 1 - Aprobado Por CECDocumento48 páginasAvance 1 - Aprobado Por CECSamuel MonzónAún no hay calificaciones
- Leptina liporregulación resistencia insulinaDocumento5 páginasLeptina liporregulación resistencia insulinaLaura Pérez RamosAún no hay calificaciones
- La Hiperglucemia Que Camino LlevaDocumento10 páginasLa Hiperglucemia Que Camino LlevaMatt AriasAún no hay calificaciones
- Diabetes FarmacologiaDocumento32 páginasDiabetes FarmacologiaLuis EstradaAún no hay calificaciones
- Taller de Fisiopatologia de La DiabetesDocumento6 páginasTaller de Fisiopatologia de La DiabetesLuisa Novoa Basto100% (1)
- Molibdeno y Sodio RemastDocumento7 páginasMolibdeno y Sodio RemastIsaac Condor MorenoAún no hay calificaciones
- BIOLOGIADocumento7 páginasBIOLOGIAJulián MelendrezAún no hay calificaciones
- Protocolo transfusión sanguíneaDocumento22 páginasProtocolo transfusión sanguíneakandy cespedes monteroAún no hay calificaciones
- OSTEOLOGÍADocumento2 páginasOSTEOLOGÍAMARVEL PEDIDOSAún no hay calificaciones
- Anatomía y Fisiología de Oído Interno UASDocumento40 páginasAnatomía y Fisiología de Oído Interno UASNicoleOrmazábal100% (1)
- Metabolismo de AminoacidosDocumento6 páginasMetabolismo de AminoacidosGaby Rodriguez ZambranoAún no hay calificaciones
- Ventilación - Mecánica de La Respiración - LecturioDocumento18 páginasVentilación - Mecánica de La Respiración - LecturioSofia Ailen IsuardiAún no hay calificaciones
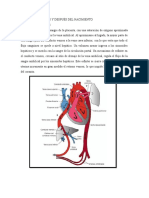
- Cambios circulatorios antes y después del nacimientoDocumento5 páginasCambios circulatorios antes y después del nacimientoDeyvid Piero Tolentino IsidroAún no hay calificaciones
- Sistema EndocrinoDocumento12 páginasSistema EndocrinoAngela SotoAún no hay calificaciones
- Fisiología del eritrocito: transporte de oxígeno y metabolismo del hierroDocumento16 páginasFisiología del eritrocito: transporte de oxígeno y metabolismo del hierroJeniAún no hay calificaciones
- HipófisisDocumento32 páginasHipófisisJossuee Paredes ContrerasAún no hay calificaciones
- Sistema de Endomembranas ComponentesDocumento8 páginasSistema de Endomembranas ComponentesLissSanchezAndradeAún no hay calificaciones
- Parascaris EquorumDocumento2 páginasParascaris Equorumpierre lizaAún no hay calificaciones
- Manejo Inicial Del Paciente Politraumatizado: Ponente: Dra. Gomez Daneila. Dr. Albahdari Jalal. Dr. Requena RayditDocumento33 páginasManejo Inicial Del Paciente Politraumatizado: Ponente: Dra. Gomez Daneila. Dr. Albahdari Jalal. Dr. Requena RayditChristian MoralesAún no hay calificaciones
- Empresa: Virrey Solis Ips S.A: Area de HematologíaDocumento2 páginasEmpresa: Virrey Solis Ips S.A: Area de HematologíaOscar ContrerasAún no hay calificaciones
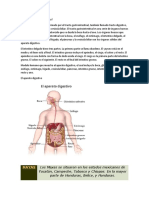
- Qué Es El Aparato DigestivoDocumento6 páginasQué Es El Aparato DigestivoMaria Tibisay NievesAún no hay calificaciones
- Personajes de La Historia de La BioquimicaDocumento6 páginasPersonajes de La Historia de La BioquimicaImanol GuzmanAún no hay calificaciones
- Farmacologia Hormona Tiroidea-1Documento70 páginasFarmacologia Hormona Tiroidea-1Edson Machicado VargasAún no hay calificaciones
- Aplicación de Vendaje Neuromuscular Como Auxiliar para TalalgiasDocumento36 páginasAplicación de Vendaje Neuromuscular Como Auxiliar para TalalgiasCarlos Antonio �Flores DelgadoAún no hay calificaciones
- Guía de Procesos Inmunológicos Primer ParcialDocumento11 páginasGuía de Procesos Inmunológicos Primer ParcialCarmen ReyesAún no hay calificaciones
- Función Renal y ExámenesDocumento29 páginasFunción Renal y ExámenesLuis Jonatan Pahuara HuamanAún no hay calificaciones
- Motivacion 01Documento15 páginasMotivacion 01Fabian MurrietaAún no hay calificaciones
- Anemia FerropénicaDocumento12 páginasAnemia FerropénicaLuu BAún no hay calificaciones
- Neurotoxicidad Af EsDocumento12 páginasNeurotoxicidad Af EsjuanAún no hay calificaciones
- Manual de Anestesia para Odontologia-Federico M. García-GodoyDocumento96 páginasManual de Anestesia para Odontologia-Federico M. García-GodoyAngie Velastegui PelaezAún no hay calificaciones
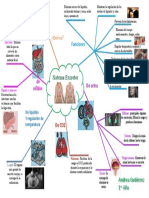
- Mapa Mental Del Sistema Excretor 1Documento1 páginaMapa Mental Del Sistema Excretor 1AndreaAún no hay calificaciones
- Amino Power - Ficha Tecnica. 20Documento3 páginasAmino Power - Ficha Tecnica. 20jaeles albAún no hay calificaciones
- Guía Examen Generalidades Medicina Fes IztacalaDocumento24 páginasGuía Examen Generalidades Medicina Fes Iztacalaguzoliver80% (5)