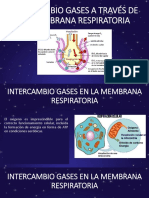
También podría gustarte
- Ley de Henry y RaoultDocumento2 páginasLey de Henry y RaoultAlvaro Michel Castillo Quispehuanca100% (2)
- Mezcla y CombinaciónDocumento5 páginasMezcla y Combinaciónmollinedo asnoAún no hay calificaciones
- Lab 10 - Equivalente de ArenaDocumento5 páginasLab 10 - Equivalente de ArenaAbiezerGonzalesMedinaAún no hay calificaciones
- Manual de MineriaDocumento158 páginasManual de MineriaJuan Carlos Castillón Vilcapoma100% (1)
- Psicometría. Principios básicos y protocolos experimentales diversosDe EverandPsicometría. Principios básicos y protocolos experimentales diversosAún no hay calificaciones
- Resumen Cap 39Documento7 páginasResumen Cap 39jonasgs_92100% (2)
- Presentación - Ley de DaltonDocumento56 páginasPresentación - Ley de DaltonDanielAún no hay calificaciones
- Ingenieria 6Documento44 páginasIngenieria 6pedroAún no hay calificaciones
- Fisiología de La Respiración.Documento42 páginasFisiología de La Respiración.carmenAún no hay calificaciones
- Sesión 1 - Fisiología Básica PDF DescargableDocumento12 páginasSesión 1 - Fisiología Básica PDF DescargableJosé Luis Torres OrtegaAún no hay calificaciones
- Apunte RESPI (1) Olor PDFDocumento35 páginasApunte RESPI (1) Olor PDFRocio DominguezAún no hay calificaciones
- Apunte de Respiratorio - RaulDocumento35 páginasApunte de Respiratorio - RaulGuilherme KuperAún no hay calificaciones
- Seminario 4Documento9 páginasSeminario 4christianrojashAún no hay calificaciones
- Fisiología Del Intercambio de GasesDocumento6 páginasFisiología Del Intercambio de GasesalbaAún no hay calificaciones
- TCP II - Parte IIDocumento18 páginasTCP II - Parte IIsebaflores.deAún no hay calificaciones
- Gasometria ArterialDocumento25 páginasGasometria ArterialErika PereraAún no hay calificaciones
- HematosisDocumento5 páginasHematosisMaría Belén MareskyAún no hay calificaciones
- Respi TotalDocumento103 páginasRespi TotalBrenda PusdáAún no hay calificaciones
- Clase 2 BIOFISICA DE GASESDocumento9 páginasClase 2 BIOFISICA DE GASESMicaela DuranAún no hay calificaciones
- 29) Transporte de Gases en Sangre, Respiración CelularDocumento12 páginas29) Transporte de Gases en Sangre, Respiración Celularyulianaledesmantvg16Aún no hay calificaciones
- Intercambio GaseosoDocumento9 páginasIntercambio GaseosoRaulAisaAún no hay calificaciones
- Aparto RespiraorioDocumento17 páginasAparto RespiraorioMilagro Ester HerreraAún no hay calificaciones
- Intercambio GaseosoDocumento26 páginasIntercambio GaseosoValeria ValladaresAún no hay calificaciones
- Capitulo 40 Fisiología de GuytonDocumento7 páginasCapitulo 40 Fisiología de Guytonwalton-miguelAún no hay calificaciones
- Clase de Intercambio Gaseoso Fisio Clase ChekDocumento2 páginasClase de Intercambio Gaseoso Fisio Clase ChekJorge Alejandro Aldana NavarroAún no hay calificaciones
- Respirator I oDocumento12 páginasRespirator I oRené MuñozAún no hay calificaciones
- Clases RespiratorioDocumento12 páginasClases RespiratorioStephanie KohlerAún no hay calificaciones
- Gases2 PDFDocumento3 páginasGases2 PDFDavid LuengasAún no hay calificaciones
- Ley de BoyleDocumento3 páginasLey de BoyleJohnGonzalezAún no hay calificaciones
- Intercambio de Gases (III)Documento18 páginasIntercambio de Gases (III)Dariana SandovalAún no hay calificaciones
- Resumen BiofisicaDocumento6 páginasResumen Biofisicamaria camila hoyosAún no hay calificaciones
- Intercambio GaseosoDocumento9 páginasIntercambio GaseosoAlex Kevin Gonzales QuispeAún no hay calificaciones
- Mezcla de Gases Biofisica Respiratorios C AudioDocumento16 páginasMezcla de Gases Biofisica Respiratorios C AudioSabrina CángeleAún no hay calificaciones
- Guía 6Documento6 páginasGuía 6Diederich JiménezAún no hay calificaciones
- Apuntes Tema 6Documento12 páginasApuntes Tema 6Ricard CrzAún no hay calificaciones
- Presión Parcial Del OxígenoDocumento9 páginasPresión Parcial Del OxígenoJosué Tanta SandovalAún no hay calificaciones
- Difusion FisiologiaDocumento6 páginasDifusion FisiologiaCarlosMS18Aún no hay calificaciones
- Tema 25Documento3 páginasTema 25gabrielmd858Aún no hay calificaciones
- Biofisica de Los GasesDocumento4 páginasBiofisica de Los GasesMalena SerranoAún no hay calificaciones
- Aspectos Fisicos Fisiol y Bioquim de La Ohb Poli 1Documento37 páginasAspectos Fisicos Fisiol y Bioquim de La Ohb Poli 1sanusdentisAún no hay calificaciones
- Biofísica de La Respiración ExternaDocumento25 páginasBiofísica de La Respiración ExternaalvaroAún no hay calificaciones
- GASESDocumento33 páginasGASESNikiMgAún no hay calificaciones
- BIOFÍSICADocumento10 páginasBIOFÍSICAcatalinarevueltasnAún no hay calificaciones
- Peso EspecificoDocumento9 páginasPeso Especifico'Aguusz Gomez'Aún no hay calificaciones
- Práctica 3. Estudio de Las Propiedades de Los Sistemas Gaseosos.Documento17 páginasPráctica 3. Estudio de Las Propiedades de Los Sistemas Gaseosos.DANIEL ENRIQUE PIÑEROS RODRIGUEZAún no hay calificaciones
- Leyes Generales de Los GasesDocumento5 páginasLeyes Generales de Los GasesBalam CaballeroAún no hay calificaciones
- Unidad 1. Respiración: Tema 1: Gases en El Aire Y en El Agua. Sistemas Respiratorios. Tipos de Sistemas RespiratoriosDocumento10 páginasUnidad 1. Respiración: Tema 1: Gases en El Aire Y en El Agua. Sistemas Respiratorios. Tipos de Sistemas RespiratoriosClara Muñoz PartidaAún no hay calificaciones
- Repaso Fisilogia e Inmunologia Semana SantaDocumento7 páginasRepaso Fisilogia e Inmunologia Semana SantaJuan Pablo Zuluaga CastañoAún no hay calificaciones
- BF DE LA RESPIRACIÒN 2024Documento26 páginasBF DE LA RESPIRACIÒN 2024helenloor81Aún no hay calificaciones
- Introduccion A La NeumologiaDocumento8 páginasIntroduccion A La NeumologiaCarlos Richard Ambrosio PiantoAún no hay calificaciones
- Fisiologia Del Intercambio de GasesDocumento18 páginasFisiologia Del Intercambio de Gasesmanuel herradoraAún no hay calificaciones
- Fisio PulmonarDocumento12 páginasFisio PulmonarClaudia Yanireth Martinez RamirezAún no hay calificaciones
- LLLDocumento11 páginasLLLurgazuAún no hay calificaciones
- Teoria Leyes de Los Gases Marzo 2024 Medicina BecadosDocumento14 páginasTeoria Leyes de Los Gases Marzo 2024 Medicina BecadosIvanAún no hay calificaciones
- Transporte de Oxigeno. Aparato Respiratorio, Fisiologia VeterinariaDocumento6 páginasTransporte de Oxigeno. Aparato Respiratorio, Fisiologia VeterinariaAndrea MendozaAún no hay calificaciones
- 13-Operación AbsorciónDocumento34 páginas13-Operación AbsorciónMiguel Antonio Soto González100% (1)
- Estado GaseosoDocumento35 páginasEstado GaseosoJohn ChinchonAún no hay calificaciones
- El Estado Gaseoso y Fuerzas IntermolecularesDocumento67 páginasEl Estado Gaseoso y Fuerzas IntermolecularesLuis Agudelo0% (1)
- Capitulo 40Documento4 páginasCapitulo 40Mariana VásquezAún no hay calificaciones
- TP Respiración Física 2015 CompletoDocumento27 páginasTP Respiración Física 2015 CompletoyolandaAún no hay calificaciones
- Unidades de Química Ii TrnnasformadoDocumento133 páginasUnidades de Química Ii TrnnasformadoDaniela BarrionuevoAún no hay calificaciones
- Gases Vapores y PsicrometriaDocumento4 páginasGases Vapores y PsicrometriaRicardo SierraAún no hay calificaciones
- Resumen Cap 40 de Guyton Principios Fisicos Del Intercambio Gaseoso - Doc 20240421 153018 0000Documento7 páginasResumen Cap 40 de Guyton Principios Fisicos Del Intercambio Gaseoso - Doc 20240421 153018 0000Aurora MéndezAún no hay calificaciones
- Gases en La Respiración PDFDocumento8 páginasGases en La Respiración PDFdexterMIAún no hay calificaciones
- Propiedades Físicas y Comportamiento Del SueloDocumento42 páginasPropiedades Físicas y Comportamiento Del SueloAlexis Julián ZuritaAún no hay calificaciones
- Conformados LadrillosDocumento4 páginasConformados LadrillosgarratemucoAún no hay calificaciones
- Alcoholemia Por El Metodo de FelsteinDocumento4 páginasAlcoholemia Por El Metodo de FelsteinSteeven RincónAún no hay calificaciones
- Manual de Patologia GeneralDocumento114 páginasManual de Patologia GeneralCarolina PavezAún no hay calificaciones
- U3 - Termodinámica de Los Seres Vivos - Física e Introducción A La BiofísicaDocumento12 páginasU3 - Termodinámica de Los Seres Vivos - Física e Introducción A La Biofísicavaleria villca medinaAún no hay calificaciones
- Power Point Generalidades de La QuímicaDocumento13 páginasPower Point Generalidades de La Químicayolima vivasAún no hay calificaciones
- Rectificación en Torres de PlatosDocumento78 páginasRectificación en Torres de PlatosAngelo SuperTello100% (1)
- Nuevos MaterialesDocumento54 páginasNuevos MaterialesAlejandro MirandaAún no hay calificaciones
- FosforenoDocumento4 páginasFosforenoDvid HórusAún no hay calificaciones
- Tds - 023 Filtralite 2100Documento2 páginasTds - 023 Filtralite 2100Armando CastañedaAún no hay calificaciones
- Materiales de Construcción-Tecnologia IndustrialDocumento10 páginasMateriales de Construcción-Tecnologia Industrialchap_plAún no hay calificaciones
- WtbetbrbwtbDocumento6 páginasWtbetbrbwtbJonathan Pinto MaytaAún no hay calificaciones
- Cinetica y Validacion Procesos Termicos 2013Documento101 páginasCinetica y Validacion Procesos Termicos 2013Gino Carbajal JavierAún no hay calificaciones
- TEMA 10 Microorganismos PDFDocumento18 páginasTEMA 10 Microorganismos PDFMarcos Belmar OrtizAún no hay calificaciones
- Pre Prueba de Sexto Grado CotejadaDocumento12 páginasPre Prueba de Sexto Grado Cotejadajohn0% (2)
- Quimica TrabajoDocumento3 páginasQuimica TrabajoNicol DuránAún no hay calificaciones
- Procedimiento Encapsulamiento de Aceite Vegetal Usado y Recolección de GrasasDocumento5 páginasProcedimiento Encapsulamiento de Aceite Vegetal Usado y Recolección de GrasasEdisson SanabriaAún no hay calificaciones
- Análisis Químico Cualitativo de Los Cationes Del Grupo I y IIDocumento6 páginasAnálisis Químico Cualitativo de Los Cationes Del Grupo I y IIIDARRAGA BASTIDAS LAURA DANIELAAún no hay calificaciones
- Actividad Colaborativa Paso 3Documento14 páginasActividad Colaborativa Paso 3Christian Olaya QuesadaAún no hay calificaciones
- Todo Sobre El GrafenoDocumento9 páginasTodo Sobre El GrafenoGino RojasAún no hay calificaciones
- JP 14 HomogenizaciónDocumento8 páginasJP 14 HomogenizaciónDiana Sheyla Izuisa GonzalesAún no hay calificaciones
- Rompedor de VacioDocumento8 páginasRompedor de VacioWalter DelgadoAún no hay calificaciones
- ArtículoDocumento3 páginasArtículojaugustonmAún no hay calificaciones
- Calculo de Flete de MaterialesDocumento8 páginasCalculo de Flete de MaterialesJimmy ManuelAún no hay calificaciones
- Caldera WordDocumento8 páginasCaldera WordRodrigo GuizarAún no hay calificaciones
- Residuos No PeligrososDocumento3 páginasResiduos No PeligrososLarry ZambranoAún no hay calificaciones