También podría gustarte
- Resumen-Flujograma (Glucolisis Y Ciclo de Krebs)Documento4 páginasResumen-Flujograma (Glucolisis Y Ciclo de Krebs)Edgar HuamanchumoAún no hay calificaciones
- Glucólisis 2024Documento13 páginasGlucólisis 2024Anna VazquezAún no hay calificaciones
- Tabla de GluconeogenesisDocumento3 páginasTabla de GluconeogenesisPerla MorenoAún no hay calificaciones
- Tema 3 GlucolisisDocumento33 páginasTema 3 Glucolisismarcosh777Aún no hay calificaciones
- Clase 9Documento55 páginasClase 9Giampierre VillenaAún no hay calificaciones
- Teorico 4 Metabolismo de GlucidosDocumento25 páginasTeorico 4 Metabolismo de GlucidosAdriano pompeiAún no hay calificaciones
- GLICÓLISISDocumento22 páginasGLICÓLISISSamuel Isaac Saenz ChaucaAún no hay calificaciones
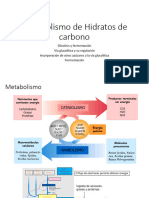
- Metabolismo de Hidratos de CarbonoDocumento9 páginasMetabolismo de Hidratos de CarbonoEmi NavarreteAún no hay calificaciones
- 1 Metabolismo de Glucidos - 3Documento61 páginas1 Metabolismo de Glucidos - 3Julio AlejandroAún no hay calificaciones
- GlucólisisDocumento10 páginasGlucólisismaria moranAún no hay calificaciones
- GluconeogenesisDocumento20 páginasGluconeogenesisdan100% (2)
- Metabolismo de Los CarbohidratosDocumento24 páginasMetabolismo de Los CarbohidratosHuaygua colque Shirley cathiaAún no hay calificaciones
- Unidades 13 y 14 Metabolismo GlucidosDocumento93 páginasUnidades 13 y 14 Metabolismo GlucidoscamilaAún no hay calificaciones
- Metabolismo de Los Carbohidratos Hasta Antes de KrebsDocumento47 páginasMetabolismo de Los Carbohidratos Hasta Antes de KrebsBrayan Stik0% (1)
- Gluconeogenesis 2.0Documento22 páginasGluconeogenesis 2.0Valeria Ake balcazarAún no hay calificaciones
- Bbiq U2 A2 AmffDocumento4 páginasBbiq U2 A2 AmffMichel FriasAún no hay calificaciones
- Iii-1-Cap 8-Glucolisis-Mckee - Parte 1Documento8 páginasIii-1-Cap 8-Glucolisis-Mckee - Parte 1Para páginasAún no hay calificaciones
- Metabolismo Anaerobio GLUCOLISISDocumento16 páginasMetabolismo Anaerobio GLUCOLISISAngieAlarcónAún no hay calificaciones
- Fermentacion OrganotrofaDocumento87 páginasFermentacion OrganotrofaKevin Soto Castillones KeVsoAún no hay calificaciones
- MetabolismoDocumento90 páginasMetabolismoDaniel TorresAún no hay calificaciones
- GlucolisisDocumento27 páginasGlucolisiskenialopez1009Aún no hay calificaciones
- Clase GlucolisisyGluconeogenesisDocumento55 páginasClase GlucolisisyGluconeogenesisAnonimo22Aún no hay calificaciones
- Temas 6-15 Parcial 2 (10 Temas)Documento75 páginasTemas 6-15 Parcial 2 (10 Temas)Ana Belén Quintanilla JuanAún no hay calificaciones
- Metabolismo de La Glucosa Parte I PDFDocumento34 páginasMetabolismo de La Glucosa Parte I PDFIsabella AcostaAún no hay calificaciones
- Via MetabólicaDocumento9 páginasVia MetabólicaEnrique OrtizAún no hay calificaciones
- Clase 9 y 10 Metabolismo de Hidratos de Carbono 2022Documento36 páginasClase 9 y 10 Metabolismo de Hidratos de Carbono 2022belencitazc2011Aún no hay calificaciones
- Clase 2 BioquimicaDocumento18 páginasClase 2 BioquimicaLeslie Rosado MerinoAún no hay calificaciones
- Ciclo de KrebsDocumento25 páginasCiclo de KrebsLuis Carlos VinuezaAún no hay calificaciones
- Fermentacion OrganotrofaDocumento86 páginasFermentacion OrganotrofaJoselyn LixAún no hay calificaciones
- Tema-4-Vision General Del Metabolismo de Glucidos.Documento36 páginasTema-4-Vision General Del Metabolismo de Glucidos.Carmen VilchesAún no hay calificaciones
- Metabolismo de CarbohidratosDocumento29 páginasMetabolismo de CarbohidratosALISSON ESTHEFANY SANTAMARIA BALAREZOAún no hay calificaciones
- Capítulo 11 GluconeogénesisDocumento3 páginasCapítulo 11 GluconeogénesisBelen Rosas MalletteAún no hay calificaciones
- Metabolismo y Función de La GlucosaDocumento10 páginasMetabolismo y Función de La GlucosaKaren MorenoAún no hay calificaciones
- Resolución Del Taller 6 - Síntesis de Glucosa, Ácidos Grasos y NADPHDocumento17 páginasResolución Del Taller 6 - Síntesis de Glucosa, Ácidos Grasos y NADPHssmrt.2016Aún no hay calificaciones
- GLUCOLISISDocumento20 páginasGLUCOLISISJefferson JerezAún no hay calificaciones
- Reporte AlimentosDocumento17 páginasReporte AlimentosSalma Montes de OcaAún no hay calificaciones
- Glicólisis, Gluconeogénesis, CK, Glucogenólisis, Glucogénesis PDFDocumento42 páginasGlicólisis, Gluconeogénesis, CK, Glucogenólisis, Glucogénesis PDF108 Alanis Anel Hernández BautistaAún no hay calificaciones
- Pentosas FosfatoDocumento9 páginasPentosas FosfatoHéctor ScandaAún no hay calificaciones
- Ejercicio Integrador Cuadro Comparativo de Rutas Metabólicas de CHO SDocumento3 páginasEjercicio Integrador Cuadro Comparativo de Rutas Metabólicas de CHO SEmiliano santosAún no hay calificaciones
- Metabolismo de Los CarbohidratosDocumento117 páginasMetabolismo de Los CarbohidratosAscrusAún no hay calificaciones
- Metabolismo de Carbohidratos - TorresButrónJesúsDocumento5 páginasMetabolismo de Carbohidratos - TorresButrónJesúsKarol MontañoAún no hay calificaciones
- Gluconeogénesis - Reacciones Enzimáticas - Dr. Alberto SanagustínDocumento5 páginasGluconeogénesis - Reacciones Enzimáticas - Dr. Alberto SanagustínJosmar Villano BonillaAún no hay calificaciones
- Formato de Entrega Tarea 4 Metabolismo Anabolismo y Catabolismo Procesos Centrales Del MetabolismoDocumento9 páginasFormato de Entrega Tarea 4 Metabolismo Anabolismo y Catabolismo Procesos Centrales Del MetabolismoJACKELINE TORRES PEDREROSAún no hay calificaciones
- GlucolisisDocumento3 páginasGlucolisismaria isidroAún no hay calificaciones
- Mapas Conceptuales Del Metabolismo de Carbohidratos 274281 Downloable 2200545Documento4 páginasMapas Conceptuales Del Metabolismo de Carbohidratos 274281 Downloable 2200545jaimeAún no hay calificaciones
- Factores Físico Químicos en La Producción Del BiogásDocumento63 páginasFactores Físico Químicos en La Producción Del BiogásPANDITA KAWAI M.Aún no hay calificaciones
- Bme U2 A2 ObvcDocumento8 páginasBme U2 A2 ObvcSilvia VelasquezAún no hay calificaciones
- Balanza Heuristica G10-ComprimidoDocumento19 páginasBalanza Heuristica G10-ComprimidoLAURA HAYDEE CASTILLO DONISAún no hay calificaciones
- 08-Semana Clase Sesià N 17 - 18 Glicolisis-Fermentaciones 2s2022Documento47 páginas08-Semana Clase Sesià N 17 - 18 Glicolisis-Fermentaciones 2s2022Luciano Bonilla ReyesAún no hay calificaciones
- Vias Metabolicas DegradativasDocumento68 páginasVias Metabolicas DegradativasRodrigo JavierAún no hay calificaciones
- Metabolismo de CarboidratosDocumento23 páginasMetabolismo de CarboidratosVerónica VelásquezAún no hay calificaciones
- Metabolismo IntermedioDocumento15 páginasMetabolismo IntermedioAdrian LeónAún no hay calificaciones
- Glucolisis 21645 PDFDocumento77 páginasGlucolisis 21645 PDFShärlÿ ContrerasAún no hay calificaciones
- Metabolismo de Los Carbohidratos y Su Relación BioenergéticaDocumento40 páginasMetabolismo de Los Carbohidratos y Su Relación BioenergéticaAnthony Xavier Tlv Ponce García100% (1)
- GluconeogénesisDocumento10 páginasGluconeogénesisSandra RiveraAún no hay calificaciones
- Síntesis Del Almidón, Lactosa y Sacarosa - Grupo 03Documento38 páginasSíntesis Del Almidón, Lactosa y Sacarosa - Grupo 03karenyamilezvAún no hay calificaciones
- Gli Colis IsDocumento26 páginasGli Colis Isvalerialarrarte01Aún no hay calificaciones
- Rutas MetabolicasDocumento9 páginasRutas MetabolicasXoana CuencaAún no hay calificaciones
- Ecuador DentalDocumento23 páginasEcuador DentalNatalia Rojas33% (3)
- Justificación Del Uso Del Cuaderno para 3 A PreescolarDocumento5 páginasJustificación Del Uso Del Cuaderno para 3 A PreescolarCaro Maya100% (1)
- Laboratorio de Ensayos IES LM-79 LM-80 LM-82 - TURCLABDocumento2 páginasLaboratorio de Ensayos IES LM-79 LM-80 LM-82 - TURCLABDario ManobandaAún no hay calificaciones
- Anexo 3 Proyectos Seleccionados 0Documento20 páginasAnexo 3 Proyectos Seleccionados 0Cesar FernandezAún no hay calificaciones
- AchicoriaDocumento14 páginasAchicoriaCesarCumbreraAún no hay calificaciones
- Revisar Entrega de Examen - Semana 6 - Sumativa 2 - DISPDocumento8 páginasRevisar Entrega de Examen - Semana 6 - Sumativa 2 - DISPecornejokAún no hay calificaciones
- AGENDA EstadNegocios JulSep2013 BANDocumento9 páginasAGENDA EstadNegocios JulSep2013 BANRa LauAún no hay calificaciones
- MANUAL DE PRACTICAS PROFESIONALES (Versión Consejo Académico) 10-03-2021Documento14 páginasMANUAL DE PRACTICAS PROFESIONALES (Versión Consejo Académico) 10-03-2021Luis MendezAún no hay calificaciones
- Script Conferencia Perfectos DragonesDocumento11 páginasScript Conferencia Perfectos DragonesdibuyoAún no hay calificaciones
- Clase - Metabolismo Fructosa, Etanol, Galactosa USSDocumento6 páginasClase - Metabolismo Fructosa, Etanol, Galactosa USSLohengrinAún no hay calificaciones
- Formato Aceptación Acuerdo SISPAE ResponsableDocumento2 páginasFormato Aceptación Acuerdo SISPAE ResponsableAngie Dayanne LeónAún no hay calificaciones
- Entidades Certificadora Vistos Buenos y UAPDocumento14 páginasEntidades Certificadora Vistos Buenos y UAPnegociosinter23164167% (6)
- Principio de OrganizaciónDocumento4 páginasPrincipio de OrganizaciónMichelle CamayoAún no hay calificaciones
- Póliza Endoso (2) - 1-11Documento11 páginasPóliza Endoso (2) - 1-11JORDAN JOSE LLICA AROCUTIPAAún no hay calificaciones
- BEFRAGUEDocumento2 páginasBEFRAGUEMateo Camilo AstudilloAún no hay calificaciones
- Rosa Francisca Ibáñez-Alfaro: CorrespondenciaDocumento19 páginasRosa Francisca Ibáñez-Alfaro: CorrespondenciaEumar AntillanoAún no hay calificaciones
- Correo - Javier Andres Alba Navarro - OutlookDocumento2 páginasCorreo - Javier Andres Alba Navarro - Outlookestefania herreraAún no hay calificaciones
- El Fichaje IsarDocumento25 páginasEl Fichaje IsarJhim Brandon Hidalgo PacayaAún no hay calificaciones
- Comentario de La Novela de J. EstevezDocumento4 páginasComentario de La Novela de J. EstevezHugo UltraAún no hay calificaciones
- Gran Manifiesto Gnostico 1971 PDFDocumento16 páginasGran Manifiesto Gnostico 1971 PDFHardy Enrique Pennekamp CantillanaAún no hay calificaciones
- Trabajo Lista de Chequeo Ferreteria Segunda EntregaDocumento54 páginasTrabajo Lista de Chequeo Ferreteria Segunda EntregaKaritho LizethAún no hay calificaciones
- Deficiencia de Vitamina B12 Como Etiología de Deterioro Cognitivo y DemenciaDocumento15 páginasDeficiencia de Vitamina B12 Como Etiología de Deterioro Cognitivo y DemenciaEricka FelizAún no hay calificaciones
- Ogunda MafunDocumento3 páginasOgunda MafunbobdiazfrancoAún no hay calificaciones
- El Proyecto VenusDocumento1 páginaEl Proyecto VenusAnonymous ko3kUoAún no hay calificaciones
- DOC1Documento5 páginasDOC1Guillermo Marcelo Badillo MuñozAún no hay calificaciones
- Dipticos GeogebraDocumento2 páginasDipticos GeogebraElizabeth ValladaresAún no hay calificaciones
- Trabajo1GOPSMBA33AGrupo2 PacificoVidav6Documento16 páginasTrabajo1GOPSMBA33AGrupo2 PacificoVidav6Atik NaiAún no hay calificaciones
- Caso ClinicoDocumento7 páginasCaso ClinicoFátima BohorquezAún no hay calificaciones
- Analisis CombinatorioDocumento7 páginasAnalisis CombinatorioRonald Mamani MaytaAún no hay calificaciones
- Resolucion SicDocumento84 páginasResolucion SicCamilo García SarmientoAún no hay calificaciones