También podría gustarte
- 5077-Texto Del Artículo-40487-1-10-20220524Documento10 páginas5077-Texto Del Artículo-40487-1-10-20220524ANGEL RAMON MACIAS RODRIGUEZAún no hay calificaciones
- Coliformes Fecales Agua-Sedimentos PDFDocumento10 páginasColiformes Fecales Agua-Sedimentos PDFCarlos MuñozAún no hay calificaciones
- Biorremediación Efluentes Camaron Utilizando Tapetes MicrobianosDocumento15 páginasBiorremediación Efluentes Camaron Utilizando Tapetes MicrobianosMónica GonzálezAún no hay calificaciones
- Innotec 1688-3691: IssnDocumento15 páginasInnotec 1688-3691: IssnSó Maquinas PesadasAún no hay calificaciones
- Article ReviewDocumento8 páginasArticle Reviewjeric gonzalezAún no hay calificaciones
- Parásitos Protozoarios EntéricosDocumento7 páginasParásitos Protozoarios Entéricoslourdes cueto sotoAún no hay calificaciones
- Identificación de Las Especies de Algas en La Represa de San José de Uzuña, Arequipa, Perú-2019Documento9 páginasIdentificación de Las Especies de Algas en La Represa de San José de Uzuña, Arequipa, Perú-2019Liseth CcosccoAún no hay calificaciones
- Contaminación Fecal en Agua PotableDocumento10 páginasContaminación Fecal en Agua PotableDane Bautista TellezAún no hay calificaciones
- V 23 N 3 A 1Documento12 páginasV 23 N 3 A 1Katherin RendonAún no hay calificaciones
- Descomposición de La Hojarasca y Su Relación Con Los MacroinvertebradosDocumento16 páginasDescomposición de La Hojarasca y Su Relación Con Los MacroinvertebradosAna Belen RobledoAún no hay calificaciones
- Microbiologia Trabajo de InvestigacionDocumento20 páginasMicrobiologia Trabajo de InvestigacionYulenyAún no hay calificaciones
- Arsenico ToacasoDocumento15 páginasArsenico ToacasoANGELA GABRIELA VERDESOTO SALAZARAún no hay calificaciones
- Diatomeas Como BioindicadoresDocumento21 páginasDiatomeas Como Bioindicadoressebastian floresAún no hay calificaciones
- Revista Cubana de Higiene y Epidemiología 0253-1751: Issn: Pmasb@infomed - Sld.cuDocumento7 páginasRevista Cubana de Higiene y Epidemiología 0253-1751: Issn: Pmasb@infomed - Sld.cuADRIANA BRIGITTE VALDIVIA ARENASAún no hay calificaciones
- Modelo Informe LabDocumento9 páginasModelo Informe LabNanis BarretoAún no hay calificaciones
- Identificación de Polimorfismos en El Gen RXAM1 de Yuca y Su Asociación Con La Resistencia A La Bacteriosis VascularDocumento9 páginasIdentificación de Polimorfismos en El Gen RXAM1 de Yuca y Su Asociación Con La Resistencia A La Bacteriosis VascularCARLOS GABRIEL DE LEON OSORIOAún no hay calificaciones
- Ecoepidemiología de Los Escorpiones de Importancia Medica en ColombiaDocumento12 páginasEcoepidemiología de Los Escorpiones de Importancia Medica en ColombiaDiego GomezAún no hay calificaciones
- C9-Microorganismos de Ecosistemas Acuáticos-MaDocumento20 páginasC9-Microorganismos de Ecosistemas Acuáticos-MaVEGA MACAS PIERO JUNIORAún no hay calificaciones
- Factores Que Favorecen La Generaci - N y Crecimiento de Algas PDFDocumento11 páginasFactores Que Favorecen La Generaci - N y Crecimiento de Algas PDFMexichem Colombia S.A.SAún no hay calificaciones
- Resumen Protocolo de Investigacion.Documento4 páginasResumen Protocolo de Investigacion.yaneliAún no hay calificaciones
- 7240-Texto Del Artículo-20361-1-10-20140409Documento9 páginas7240-Texto Del Artículo-20361-1-10-20140409Yaremi Mejia GuerreroAún no hay calificaciones
- Efecto de La Contaminacion Por Hidrocarburos Sobre Propiedades Quimicas y Microbiologicas de Un SueloDocumento9 páginasEfecto de La Contaminacion Por Hidrocarburos Sobre Propiedades Quimicas y Microbiologicas de Un Suelofanny ruiz de gomezAún no hay calificaciones
- Primer Examen de BiologíaDocumento41 páginasPrimer Examen de BiologíaKay MoralesAún no hay calificaciones
- Tarea 3. Comunidades MicrobianasDocumento6 páginasTarea 3. Comunidades MicrobianasrosmiraAún no hay calificaciones
- Metagenomic 2019Documento24 páginasMetagenomic 2019Arturo LiñanAún no hay calificaciones
- Estudio Microbiológico Del Agua en La AmidiqDocumento14 páginasEstudio Microbiológico Del Agua en La AmidiqJavier Daniel Ramírez AmayaAún no hay calificaciones
- Sandoval 2009 ADocumento9 páginasSandoval 2009 AGerardo AngoaAún no hay calificaciones
- 097 MNMNNDocumento74 páginas097 MNMNNJhoan SebastianAún no hay calificaciones
- 10881-Texto Del Articulo-32802-2-10-20210420Documento3 páginas10881-Texto Del Articulo-32802-2-10-20210420Tf ElbacanAún no hay calificaciones
- Cultivo Esponjas Venezuela PDFDocumento12 páginasCultivo Esponjas Venezuela PDFCAROLINAAún no hay calificaciones
- Articulo Original Bacterias de AguaDocumento25 páginasArticulo Original Bacterias de Aguailfeguar95Aún no hay calificaciones
- Diatomeas Como BioindicadoresDocumento21 páginasDiatomeas Como BioindicadoresMónica Nina InchuñaAún no hay calificaciones
- PDFDocumento12 páginasPDFHenry arias mAún no hay calificaciones
- Mendoza CLDocumento323 páginasMendoza CLcarlosAún no hay calificaciones
- 950 1498 1 SMDocumento7 páginas950 1498 1 SMIrene SilvaAún no hay calificaciones
- Manual de Ejercitación 2012Documento77 páginasManual de Ejercitación 2012Angelina SandovalAún no hay calificaciones
- Profesor Jose Araujo Resumenes de Seminario de Microbiologia Ambiental IIDocumento6 páginasProfesor Jose Araujo Resumenes de Seminario de Microbiologia Ambiental IIyarubitAún no hay calificaciones
- La Calidad Del Agua, en La Laguna de Tres Palos, Municipio deDocumento4 páginasLa Calidad Del Agua, en La Laguna de Tres Palos, Municipio deDylan RochaAún no hay calificaciones
- Diversidad de MexicoDocumento201 páginasDiversidad de MexicoLucia LopezAún no hay calificaciones
- Bacteriología y FitoplanctonDocumento11 páginasBacteriología y FitoplanctonRoiser Jadir Torres MarquezAún no hay calificaciones
- Metodologia para Evaluacion de Los Macroinvertbrados Acuaticos Como Indicadores de Los RecursosDocumento139 páginasMetodologia para Evaluacion de Los Macroinvertbrados Acuaticos Como Indicadores de Los RecursosDavid Arturo Pinilla OlarteAún no hay calificaciones
- Proy - Inves - 18I - PyERN (Germán Castro Mejía)Documento22 páginasProy - Inves - 18I - PyERN (Germán Castro Mejía)miguel angel muedano sanchezAún no hay calificaciones
- Metodos de Ecologia IIDocumento16 páginasMetodos de Ecologia IIRosse Elizabeth LazarteAún no hay calificaciones
- Caño Zuria Paper FinalDocumento15 páginasCaño Zuria Paper FinalAlejandra OrtizAún no hay calificaciones
- Acta Cientíifica Venezolana, Número 55, Suplemento 1, Año 2004. AsoVACDocumento278 páginasActa Cientíifica Venezolana, Número 55, Suplemento 1, Año 2004. AsoVACLuis Eduardo Traviezo VallesAún no hay calificaciones
- HeliothisDocumento6 páginasHeliothisEslymAún no hay calificaciones
- Articulo Publicado Calidad Bacteriológica de Las Piscinas de La Ciudad de Jaén, 2022 - Ciencia Latina Revista Científica MultidisciplinarDocumento13 páginasArticulo Publicado Calidad Bacteriológica de Las Piscinas de La Ciudad de Jaén, 2022 - Ciencia Latina Revista Científica MultidisciplinarALEXANDER ALDAIR PEREZ NAVARROAún no hay calificaciones
- PROYECTO DE AULA AyerDocumento19 páginasPROYECTO DE AULA AyerSaray BuelvasAún no hay calificaciones
- Lectura y Analisis de ArticulosDocumento3 páginasLectura y Analisis de ArticulosJose CumbicosAún no hay calificaciones
- Dialnet DiversidadDeMicroalgasYCianobacteriasEnMuestrasPro 6537243 PDFDocumento21 páginasDialnet DiversidadDeMicroalgasYCianobacteriasEnMuestrasPro 6537243 PDFHelcib De Los AngelesAún no hay calificaciones
- Informe 4 Presencia de Microorganismos Cultivables en Sisitemas AcuaticosDocumento6 páginasInforme 4 Presencia de Microorganismos Cultivables en Sisitemas AcuaticosPauli Martinez BendañaAún no hay calificaciones
- Catálogo de los copépodos (Crustacea: Copepoda: Calanoida y Cyclopoida) de cuerpos de agua temporales de Jalisco, México: Taxonomía y distribuciónDe EverandCatálogo de los copépodos (Crustacea: Copepoda: Calanoida y Cyclopoida) de cuerpos de agua temporales de Jalisco, México: Taxonomía y distribuciónAún no hay calificaciones
- Primera Entrega de Proyecto Escrito - Rodriguez Holguín Manuel FelipeDocumento5 páginasPrimera Entrega de Proyecto Escrito - Rodriguez Holguín Manuel Felipelaura. vargas.16Aún no hay calificaciones
- Bacterias Indicadoras de Calidad de AguaDocumento10 páginasBacterias Indicadoras de Calidad de AguaJose BarrientosAún no hay calificaciones
- FITOPLANCTONDocumento11 páginasFITOPLANCTONCristian Basilik CasasAún no hay calificaciones
- Proyecto de Micro EnterobacteriasDocumento18 páginasProyecto de Micro EnterobacteriasfiorelaAún no hay calificaciones
- 54014203Documento7 páginas54014203Jonathan MarchánAún no hay calificaciones
- Admin,+art +1Documento10 páginasAdmin,+art +1oerstedella1Aún no hay calificaciones
- Proyecto FANDocumento13 páginasProyecto FANFernando Alcivar DelgadoAún no hay calificaciones
- Contaminación de Aguas - Lectura Unidad 3Documento5 páginasContaminación de Aguas - Lectura Unidad 3Lidia Giovany Cielos AmarilloAún no hay calificaciones
- Geo U7 l173 Delta HidrografiaDocumento1 páginaGeo U7 l173 Delta HidrografiaIvanAntonioVilchezAún no hay calificaciones
- 541395S718Documento44 páginas541395S718IvanAntonioVilchezAún no hay calificaciones
- Caiman de La CostaDocumento2 páginasCaiman de La CostaIvanAntonioVilchezAún no hay calificaciones
- 2022 Tesis Neider Yesid CabreraDocumento127 páginas2022 Tesis Neider Yesid CabreraIvanAntonioVilchezAún no hay calificaciones
- Coloca Los Nombres en Su Lugar CorrespondienteDocumento1 páginaColoca Los Nombres en Su Lugar CorrespondienteIvanAntonioVilchezAún no hay calificaciones
- El Reino Monera Cianobacterias para Segundo de SecundariaDocumento5 páginasEl Reino Monera Cianobacterias para Segundo de SecundariaIvanAntonioVilchezAún no hay calificaciones
- Reino MoneraDocumento4 páginasReino MoneraIvanAntonioVilchez100% (7)
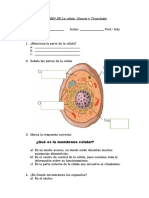
- EXAMEN DE La CélulaDocumento2 páginasEXAMEN DE La CélulaIvanAntonioVilchez100% (4)
- Ensayos Sobre MateriakDocumento25 páginasEnsayos Sobre MateriakAraceli Valladares GonzálezAún no hay calificaciones
- Grading BasicoDocumento28 páginasGrading BasicoRaul Bedoya HerediaAún no hay calificaciones
- Morfologia Interna Del CerebeloDocumento11 páginasMorfologia Interna Del CerebeloRosario M. Muñoz FloresAún no hay calificaciones
- Paletizacion de Camu CamuDocumento9 páginasPaletizacion de Camu Camuwagner alva cotrinaAún no hay calificaciones
- Determinantes para La Regeneracion de Drimys Winteri - 20221019 - 0001Documento56 páginasDeterminantes para La Regeneracion de Drimys Winteri - 20221019 - 0001olecram zelaznogAún no hay calificaciones
- Entrega 1 - Medicina PreventivaDocumento5 páginasEntrega 1 - Medicina PreventivaCAMILO CENDALESAún no hay calificaciones
- Secme 5145Documento49 páginasSecme 5145Chombo Bororow PvAún no hay calificaciones
- Voley EnsayoDocumento2 páginasVoley EnsayoYesmith Campos100% (1)
- Exposicion Salud OcupacionalDocumento6 páginasExposicion Salud OcupacionalSOFIA VALENTINA SUAREZ TORRESAún no hay calificaciones
- Diseño de Una Planta para La Producción de Compost en La Zona Norte Del Estado AnzoateguiDocumento224 páginasDiseño de Una Planta para La Producción de Compost en La Zona Norte Del Estado AnzoateguiJesus FrancoAún no hay calificaciones
- MANUAL PRÁCTICO DE Nendo-DangoDocumento54 páginasMANUAL PRÁCTICO DE Nendo-DangoNestor GuevaraAún no hay calificaciones
- Mensual OctubreDocumento2 páginasMensual OctubreLoreto Francisca Soto SalazarAún no hay calificaciones
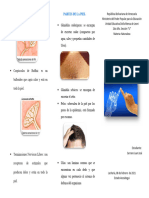
- Triptico El TactoDocumento2 páginasTriptico El Tactorafaelann711Aún no hay calificaciones
- PRACTICA 4. Descripción Morfológica y Anatómica de Flores y FrutosDocumento28 páginasPRACTICA 4. Descripción Morfológica y Anatómica de Flores y FrutosJAZMIN RUBI ALVAREZ FELIXAún no hay calificaciones
- Guia de QuimicaDocumento228 páginasGuia de QuimicaGustavo Franco Alvarez RamosAún no hay calificaciones
- Fundamentos Del Comportamiento Individual de La ComunicaciónDocumento7 páginasFundamentos Del Comportamiento Individual de La ComunicaciónLucia QuirozAún no hay calificaciones
- Huella EcologicaDocumento2 páginasHuella EcologicaJuliet LC100% (1)
- Síndrome NefróticoDocumento50 páginasSíndrome NefróticoMariajo RochaAún no hay calificaciones
- Guia de Cuantificacion CarbonoDocumento21 páginasGuia de Cuantificacion CarbonoNàpoles RaymundoAún no hay calificaciones
- 93231220-Casos-Clinicos-Automacao-Sysmex MUY BUENO PDFDocumento60 páginas93231220-Casos-Clinicos-Automacao-Sysmex MUY BUENO PDFromerilloAún no hay calificaciones
- Regla de Farrar para ApiculturaDocumento2 páginasRegla de Farrar para ApiculturaPaul Salcedo HerreraAún no hay calificaciones
- Evolucion Molecular 4Documento22 páginasEvolucion Molecular 4Gmo AnraAún no hay calificaciones
- Ataque Con Gas Sarín Al Subte de TokioDocumento10 páginasAtaque Con Gas Sarín Al Subte de TokioManusan06Aún no hay calificaciones
- Teoria de Los ImpactosDocumento7 páginasTeoria de Los ImpactosKarl A100% (1)
- A1.) Ecología, Estado Poblacional y Distribucion de La Guatusa de Roatan D. Ruatanica en Cuatro Islas Del Municipio de Roatan, Islas de La Bahia, HondurasDocumento7 páginasA1.) Ecología, Estado Poblacional y Distribucion de La Guatusa de Roatan D. Ruatanica en Cuatro Islas Del Municipio de Roatan, Islas de La Bahia, Honduraselbiciclista0% (1)
- Eco-ACTUALIZACIÓN LB BIOLÓGICA, FLORA Y FAUNA ACUÁTICADocumento83 páginasEco-ACTUALIZACIÓN LB BIOLÓGICA, FLORA Y FAUNA ACUÁTICAMarco AntonioAún no hay calificaciones
- Fche Ep 505 PDFDocumento136 páginasFche Ep 505 PDFsara rodriguezAún no hay calificaciones
- Ap Gerb-PFVC '05 PDFDocumento11 páginasAp Gerb-PFVC '05 PDFPao GabrielAún no hay calificaciones
- Fases y Etapas Fenológicas Del MaízDocumento5 páginasFases y Etapas Fenológicas Del Maízcaterine patricia inoñan yanayaco100% (1)
- Guía No. 5 de Agrícola de NovenoDocumento3 páginasGuía No. 5 de Agrícola de Novenoalejandro SandovalAún no hay calificaciones