También podría gustarte
- Sistema Nervioso Periferico ExpoDocumento8 páginasSistema Nervioso Periferico Expoivana milagros reyes acostaAún no hay calificaciones
- Comunicacion Neuroral Tema2Documento6 páginasComunicacion Neuroral Tema2Miguel Antonio NMMAún no hay calificaciones
- Vias de Conducción NerviosaDocumento9 páginasVias de Conducción NerviosaRoxanaSantillanAún no hay calificaciones
- Vias NerviosasDocumento21 páginasVias NerviosasRoxanaSantillanAún no hay calificaciones
- Fisiología de Los ReflejosDocumento10 páginasFisiología de Los ReflejosmiiaestefaniiaAún no hay calificaciones
- Practica 10 Sistema Nervioso PeriféricoDocumento9 páginasPractica 10 Sistema Nervioso PeriféricoRodolfo Garcia Urrutia MirandaAún no hay calificaciones
- Dermatomas y GangliosDocumento27 páginasDermatomas y GangliosJosé Martín J MtzAún no hay calificaciones
- 253118770-Vias-de-Conduccion-NerviosaDocumento9 páginas253118770-Vias-de-Conduccion-NerviosaJohn QuiñeAún no hay calificaciones
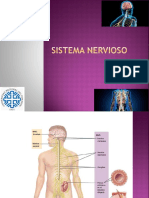
- Sistema NerviosoDocumento10 páginasSistema NerviosoLuissJQMAún no hay calificaciones
- 6histologia Del Tejido NerviosoDocumento118 páginas6histologia Del Tejido NerviosoMakarena MonsalvesAún no hay calificaciones
- Sistema NerviosoDocumento12 páginasSistema NerviosoGabriela Sofia PacAún no hay calificaciones
- Práctica Ix Tejido NerviosoDocumento13 páginasPráctica Ix Tejido NerviosoMabel AlbertoAún no hay calificaciones
- Anatomía y Fisiología del Sistema NerviosoDe EverandAnatomía y Fisiología del Sistema NerviosoCalificación: 4 de 5 estrellas4/5 (19)
- Componentes Del Sistema NerviosoDocumento6 páginasComponentes Del Sistema NerviosoJazmin Martinez zambranoAún no hay calificaciones
- 6 - Generalidades de NeurologiaDocumento12 páginas6 - Generalidades de NeurologiaFernanda de MeloAún no hay calificaciones
- Tejido NerviosoDocumento5 páginasTejido NerviosoJAVIERA FERNANDA MUÑOZAún no hay calificaciones
- Sistema Nervioso Periferico 201410Documento11 páginasSistema Nervioso Periferico 201410Edison QuembaAún no hay calificaciones
- Sistema NerviosoDocumento6 páginasSistema Nerviosoelianna matosAún no hay calificaciones
- NeurotransmisiónDocumento8 páginasNeurotransmisiónvtaglemendoza5223Aún no hay calificaciones
- Clase N°8. Sistema Nervioso PerifericoDocumento38 páginasClase N°8. Sistema Nervioso PerifericoDane Rueda0% (1)
- Tejido Nervioso HistologiaDocumento21 páginasTejido Nervioso HistologiaEliana Bell SevericheAún no hay calificaciones
- Fisiopatología Del SNDocumento35 páginasFisiopatología Del SNPaul GarciaAún no hay calificaciones
- Tejido NerviosoDocumento15 páginasTejido NerviosoEmberson Vela Gavilanes100% (1)
- Neuroanatomía y anatomía topográficaDocumento2 páginasNeuroanatomía y anatomía topográficaCelia SalmeronAún no hay calificaciones
- MORFO SOMATOSENSORIAL (1) Arti IVDocumento19 páginasMORFO SOMATOSENSORIAL (1) Arti IVMalena EcheverríaAún no hay calificaciones
- Trabajo de Investigación Sobre Sistema SomatosensorialDocumento16 páginasTrabajo de Investigación Sobre Sistema SomatosensorialAntonio LopezAún no hay calificaciones
- Practica 3 Sistema NerviosoDocumento6 páginasPractica 3 Sistema NerviosoZyprexa 17Aún no hay calificaciones
- Aparato TegumentarioDocumento12 páginasAparato TegumentarioAngeles MendozaAún no hay calificaciones
- SNC y SNPDocumento49 páginasSNC y SNPAlexisAún no hay calificaciones
- Sistema Nervioso: Estructura, Funciones y Tipos de CélulasDocumento27 páginasSistema Nervioso: Estructura, Funciones y Tipos de CélulasCatta ArellanoAún no hay calificaciones
- WernikeDocumento9 páginasWernikeElizabeth DislaAún no hay calificaciones
- Audisio E. (2014) - Transmision en El Sistema Nervioso (Ficha de Cátedra)Documento14 páginasAudisio E. (2014) - Transmision en El Sistema Nervioso (Ficha de Cátedra)Leo B BartolomeoAún no hay calificaciones
- ARCO Y ACTO REFLEJO EtcDocumento9 páginasARCO Y ACTO REFLEJO Etcjosseline riveraAún no hay calificaciones
- Apuntes de NeuroanatomiaDocumento10 páginasApuntes de NeuroanatomiaSteffany MatosAún no hay calificaciones
- Pares Craneales y CerebeloDocumento67 páginasPares Craneales y CerebeloNatalia CárdenasAún no hay calificaciones
- Resumen Sistema NerviosoDocumento12 páginasResumen Sistema NerviosoDeinir SolanoAún no hay calificaciones
- OSBGT56Documento16 páginasOSBGT56alexantonjazzAún no hay calificaciones
- Vias (Autoguardado)Documento79 páginasVias (Autoguardado)marit555Aún no hay calificaciones
- Repaso de AnatomiaDocumento8 páginasRepaso de AnatomiaEnriqueta MARTINEZAún no hay calificaciones
- 2 - Fundamentos Biológicos de La ConductaDocumento8 páginas2 - Fundamentos Biológicos de La ConductaantonioAún no hay calificaciones
- AnatomiaDocumento3 páginasAnatomiaClau FloresAún no hay calificaciones
- Practica #7 Sistema Nervioso ValpdfDocumento18 páginasPractica #7 Sistema Nervioso ValpdfValeria OrtizAún no hay calificaciones
- Tejido Nervioso1Documento19 páginasTejido Nervioso1Mathías Gonzalez MoraAún no hay calificaciones
- Resumen Final - BASES IDocumento39 páginasResumen Final - BASES IGimena Pellini GaynorAún no hay calificaciones
- Sistema NerviosoDocumento9 páginasSistema NerviosoEDGAR ESPINOZAAún no hay calificaciones
- Medula y ReflejosDocumento34 páginasMedula y ReflejosWendy JimenezAún no hay calificaciones
- Sistema nervioso y respiratorio en deportesDocumento20 páginasSistema nervioso y respiratorio en deportesDavid Quispe Jimenez100% (1)
- Nervios RaquídeosDocumento14 páginasNervios RaquídeosJuniorEspejoAún no hay calificaciones
- Sistema NerviosoDocumento20 páginasSistema NerviosoAndrea L. CernaAún no hay calificaciones
- Anatomia y Fisiologia Del Sistema NervioDocumento48 páginasAnatomia y Fisiologia Del Sistema NervioJose MaldonadoAún no hay calificaciones
- Anatomía Medula EspinalDocumento4 páginasAnatomía Medula EspinalIvania TorresAún no hay calificaciones
- Fisiología Del TactoDocumento3 páginasFisiología Del TactoAnabel IbañezAún no hay calificaciones
- Sistema Nervioso Autónomo o VegetativoDocumento12 páginasSistema Nervioso Autónomo o VegetativoCyber CyberAún no hay calificaciones
- Sistema SomatosensorialDocumento111 páginasSistema SomatosensorialDiana OlivaresAún no hay calificaciones
- P 9 Szmmdfsda &expires 1215573344&awsaccesskeyid 1 Z 5 T 9 H 8 PQ 39 V 6 F 79 V 8 G 2Documento56 páginasP 9 Szmmdfsda &expires 1215573344&awsaccesskeyid 1 Z 5 T 9 H 8 PQ 39 V 6 F 79 V 8 G 2api-3735753Aún no hay calificaciones
- Monografía - Vías Sensitivas SuperficialesDocumento12 páginasMonografía - Vías Sensitivas Superficialesleonardo.riberoAún no hay calificaciones
- Fibras NerviosasDocumento8 páginasFibras NerviosasErika VelásquezAún no hay calificaciones
- Resumen Del Sistema Nervioso AutónomoDocumento8 páginasResumen Del Sistema Nervioso AutónomoNadiaUwUAún no hay calificaciones
- Carpeta Grupal de BiologiaDocumento7 páginasCarpeta Grupal de BiologiaDaisy BscAún no hay calificaciones
- Sist AutonomoDocumento16 páginasSist AutonomoSofíaAún no hay calificaciones
- Demostración de la actividad y inhibición de la citocromo oxidasaDocumento12 páginasDemostración de la actividad y inhibición de la citocromo oxidasaNancy Membreño GaldamezAún no hay calificaciones
- SistemanerviosoautonomoDocumento39 páginasSistemanerviosoautonomoAndré FelicesAún no hay calificaciones
- Bar CP 11 Micro FDGDFGDDDocumento21 páginasBar CP 11 Micro FDGDFGDDNancy Membreño GaldamezAún no hay calificaciones
- Histologia y Embriologia Del Sistema Nervioso.: Clase Practica #4Documento8 páginasHistologia y Embriologia Del Sistema Nervioso.: Clase Practica #4Nancy Membreño GaldamezAún no hay calificaciones
- 5 Mi PreguntaDocumento3 páginas5 Mi PreguntaNancy Membreño GaldamezAún no hay calificaciones
- Análisis fisiológico de la visiónDocumento3 páginasAnálisis fisiológico de la visiónNancy Membreño GaldamezAún no hay calificaciones
- Clase 1 BioquimicaDocumento6 páginasClase 1 BioquimicaNancy Membreño GaldamezAún no hay calificaciones
- Lab 8Documento36 páginasLab 8Nancy Membreño GaldamezAún no hay calificaciones
- Examen 1 Enero 2018 Preguntas y RespuestasDocumento9 páginasExamen 1 Enero 2018 Preguntas y RespuestasNancy Membreño GaldamezAún no hay calificaciones
- Guía Plan de Procesamiento, Presentación y Análisis de DatosDocumento2 páginasGuía Plan de Procesamiento, Presentación y Análisis de DatosNancy Membreño GaldamezAún no hay calificaciones
- Siglas de Las Figuras de BarrDocumento7 páginasSiglas de Las Figuras de BarrNancy Membreño GaldamezAún no hay calificaciones
- Practico 2 MicroDocumento110 páginasPractico 2 MicroNancy Membreño GaldamezAún no hay calificaciones
- Laboratorio N. 8: Organización Interna Del Tallo Encefálico. Grandes Vías Ascendentes y DescendentesDocumento5 páginasLaboratorio N. 8: Organización Interna Del Tallo Encefálico. Grandes Vías Ascendentes y DescendentesNancy Membreño GaldamezAún no hay calificaciones
- Ilovepdf MergedDocumento82 páginasIlovepdf MergedNancy Membreño GaldamezAún no hay calificaciones
- Exámen RepetidoDocumento9 páginasExámen RepetidoNancy Membreño GaldamezAún no hay calificaciones
- PesiDocumento1 páginaPesiNancy Membreño GaldamezAún no hay calificaciones
- Operalización de VariablesDocumento1 páginaOperalización de VariablesNancy Membreño GaldamezAún no hay calificaciones
- Paradigmas ModernosDocumento41 páginasParadigmas ModernosNancy Membreño GaldamezAún no hay calificaciones
- Tema Comu SalvacionDocumento1 páginaTema Comu SalvacionNancy Membreño GaldamezAún no hay calificaciones
- Discusión 03 Objetivos 1,2,3Documento9 páginasDiscusión 03 Objetivos 1,2,3Nancy Membreño GaldamezAún no hay calificaciones
- Sistema MuscularDocumento4 páginasSistema MuscularmiguelAún no hay calificaciones
- Atención Del Parto NormalDocumento34 páginasAtención Del Parto NormalValeria Paola Molina PeinadoAún no hay calificaciones
- Triptico Mitosis-MeiosisDocumento2 páginasTriptico Mitosis-Meiosiseddy noriegaAún no hay calificaciones
- VacutainerDocumento3 páginasVacutainerAhtziri MartinezAún no hay calificaciones
- FISIOLOGÍA DEL PÁNCREAS Y SUS FUNCIONES ENDOCRINAS Y EXOCRINASDocumento6 páginasFISIOLOGÍA DEL PÁNCREAS Y SUS FUNCIONES ENDOCRINAS Y EXOCRINASRaúl R. CeledónAún no hay calificaciones
- La Investigación Con Células MadresDocumento5 páginasLa Investigación Con Células MadresYuichi MiyanowakiAún no hay calificaciones
- Técnicas histológicas microscopiaDocumento123 páginasTécnicas histológicas microscopiaKiara TejadaAún no hay calificaciones
- Fisiologia de La AudicionDocumento11 páginasFisiologia de La AudicionNitgma DcAún no hay calificaciones
- Sistema Muscular ForoDocumento6 páginasSistema Muscular ForoRony ChicoAún no hay calificaciones
- Sistema Digestivo y RespiratorioDocumento7 páginasSistema Digestivo y RespiratorioViviana OrdoñezAún no hay calificaciones
- Pseudocelomados y CelomadosDocumento38 páginasPseudocelomados y Celomadosjose carlosAún no hay calificaciones
- Tríptico Tejido MuscularDocumento2 páginasTríptico Tejido MuscularLorena PGAún no hay calificaciones
- Guía 4 Biología Del Desarrollo PDFDocumento4 páginasGuía 4 Biología Del Desarrollo PDFAngie CeronAún no hay calificaciones
- Sistema renal fisiologíaDocumento15 páginasSistema renal fisiologíaOlga Lucia Hernandez HincapieAún no hay calificaciones
- Informe 1 Histología PrácticaDocumento11 páginasInforme 1 Histología PrácticaEmery Sofía C. D. BustamanteAún no hay calificaciones
- La Piel y Las Membranas CroporalesDocumento12 páginasLa Piel y Las Membranas CroporalesAriadna La Rosa FernàndezAún no hay calificaciones
- Medula espinal corte transversalDocumento14 páginasMedula espinal corte transversalVioleta Aguayo MüllerAún no hay calificaciones
- Vagina, Utero AnexosDocumento18 páginasVagina, Utero Anexoscatastrofe1379Aún no hay calificaciones
- Examen Práctico Morfofunción I 2020 - TM-MORFOFUNCIÓN IDocumento24 páginasExamen Práctico Morfofunción I 2020 - TM-MORFOFUNCIÓN Iconche tumadreAún no hay calificaciones
- Sistema Endocrino 1Documento1 páginaSistema Endocrino 1Yesenia Mariluz Flores Flores100% (1)
- Anatomía y Fisiología de La Reproducción en Animales DomesticosDocumento63 páginasAnatomía y Fisiología de La Reproducción en Animales DomesticosPily Gato100% (3)
- Resumen Capítulo 2-Amebiasis IntestinalDocumento4 páginasResumen Capítulo 2-Amebiasis IntestinalLilly Sanmartín100% (1)
- Acrostico 1ro 86Documento2 páginasAcrostico 1ro 86Nico NichiAún no hay calificaciones
- Generalidades Del Examen FisicoDocumento48 páginasGeneralidades Del Examen FisicoJuan Guillermo Pinzón Rodríguez100% (1)
- Informe Sistema DigestivoDocumento7 páginasInforme Sistema DigestivoJose Luis PilcoAún no hay calificaciones
- Fisiologia Del PartoDocumento12 páginasFisiologia Del PartoOvidio Reinoza0% (1)
- Infografía 10 Tips Home Office Moderno Estructurado Turquesa NaranjaDocumento1 páginaInfografía 10 Tips Home Office Moderno Estructurado Turquesa NaranjaDavid Omar HernandezAún no hay calificaciones
- Ambiguedad de SexoDocumento29 páginasAmbiguedad de SexoIneska Bermudez seijasAún no hay calificaciones
- Crecimiento y Desarrollo Humano 1 (3197)Documento92 páginasCrecimiento y Desarrollo Humano 1 (3197)Luz HuamanAún no hay calificaciones
- El Ciclo de Vida de Una RanaDocumento3 páginasEl Ciclo de Vida de Una RanaGUILLERMO MURGA MASAún no hay calificaciones