También podría gustarte
- Principios básicos de patología para fisioterapia: Volumen IDe EverandPrincipios básicos de patología para fisioterapia: Volumen IAún no hay calificaciones
- Ingeniería GenéticaDocumento7 páginasIngeniería GenéticaAlejandro ZambranoAún no hay calificaciones
- Respuesta Metabolic A Al Trauma SchwartzDocumento22 páginasRespuesta Metabolic A Al Trauma SchwartzPatrick Rodas63% (8)
- MielopoyesisDocumento17 páginasMielopoyesisRodrigo Fernando Fernandez ApazaAún no hay calificaciones
- Filogenia Y Ontogenia Del Sistema NerviosoDocumento5 páginasFilogenia Y Ontogenia Del Sistema NerviosoCamila Garcia OrozcoAún no hay calificaciones
- Prueba de Ciencias Naturales 7 Unidad 2Documento3 páginasPrueba de Ciencias Naturales 7 Unidad 2Viviana Kusch Monje100% (1)
- Sesion de Cy T 2° EL SISTEMA DIGESTIVODocumento12 páginasSesion de Cy T 2° EL SISTEMA DIGESTIVOMaria FernandaAún no hay calificaciones
- MartinezFeliciano Maricela M16S4PIDocumento8 páginasMartinezFeliciano Maricela M16S4PImary martinez100% (1)
- Art 50 362Documento6 páginasArt 50 362Flor Elena ToxquiAún no hay calificaciones
- Practica #2Documento3 páginasPractica #2Alexander ZevallosAún no hay calificaciones
- Familia Hedgehog 2Documento17 páginasFamilia Hedgehog 2De La Torre Corral UrielAún no hay calificaciones
- SEMANA 9 Clase 17. Embriología IDocumento60 páginasSEMANA 9 Clase 17. Embriología IPathi VargasAún no hay calificaciones
- Genes HoxDocumento9 páginasGenes HoxGrayFullbster451Aún no hay calificaciones
- Evolución y DesarrolloDocumento34 páginasEvolución y Desarrolloagustinagonzalez525Aún no hay calificaciones
- MFPH I Semana 12 Final Orientac Estudi y Prof LESVIA URBINADocumento9 páginasMFPH I Semana 12 Final Orientac Estudi y Prof LESVIA URBINAIkuto Tsukiyomi UchihaAún no hay calificaciones
- NticaDocumento72 páginasNticacarlos Johanson Tut MoralesAún no hay calificaciones
- Holobiontes Como Unidades EvolutivasDocumento2 páginasHolobiontes Como Unidades EvolutivasJose ParraguezAún no hay calificaciones
- Ensayo de Fisiologia t5Documento8 páginasEnsayo de Fisiologia t5Ruben One31Aún no hay calificaciones
- Neurospora CrassaDocumento3 páginasNeurospora CrassaQue te importaAún no hay calificaciones
- Generación de Animales Transgénicos.Documento7 páginasGeneración de Animales Transgénicos.Fernanda SilvaAún no hay calificaciones
- Fichas de HerenciaYgeneticaDocumento4 páginasFichas de HerenciaYgeneticaofeAún no hay calificaciones
- Genetica y Psicologia El Nuevo ParadigmaDocumento11 páginasGenetica y Psicologia El Nuevo Paradigmaapi-3699471100% (3)
- Tarea # 2 Aproximacion Al Estudiio Del Comportamiento AnimalDocumento5 páginasTarea # 2 Aproximacion Al Estudiio Del Comportamiento AnimalJohanny RamirezAún no hay calificaciones
- Seccion 1 de Microbiologia Regencia OkDocumento19 páginasSeccion 1 de Microbiologia Regencia OkEdwin Emir Silva MartinezAún no hay calificaciones
- Genética de Hongos - 1Documento10 páginasGenética de Hongos - 1Gedeon CastilloAún no hay calificaciones
- Biotipos Fenotipos Genotipos (ZERON RMP 2010)Documento8 páginasBiotipos Fenotipos Genotipos (ZERON RMP 2010)Agustin ZeronAún no hay calificaciones
- Genética de Hongos OriginalDocumento17 páginasGenética de Hongos OriginalGedeon Castillo100% (1)
- Tema 1: Introducción A La Genética GenéticaDocumento167 páginasTema 1: Introducción A La Genética GenéticaMelisa BarbasAún no hay calificaciones
- Ihh Indian HedgehogDocumento4 páginasIhh Indian HedgehogDaniel Londoño100% (1)
- Libro Mejoramiento Genetico 160302051610Documento191 páginasLibro Mejoramiento Genetico 160302051610NERO3000Aún no hay calificaciones
- Unidad 3 Tarea 5 GP 151009 120Documento15 páginasUnidad 3 Tarea 5 GP 151009 120Torcoroma CarrascalAún no hay calificaciones
- HelmintosDocumento4 páginasHelmintosAlejandro BGAún no hay calificaciones
- Redolar CompletoDocumento479 páginasRedolar CompletoCarenSebastianNovellAún no hay calificaciones
- Que Es La GeneticaDocumento13 páginasQue Es La GeneticaJose Armando PallaresAún no hay calificaciones
- 4TA A 8VA SEMANA A PDFDocumento12 páginas4TA A 8VA SEMANA A PDFMaryori Treyci ILasaca ValenzuelaAún no hay calificaciones
- Primer Guia de GeneticaDocumento3 páginasPrimer Guia de GeneticaErick RamirezAún no hay calificaciones
- Balotario de Derecho Genetico-Eduardo Ayala TandazoDocumento5 páginasBalotario de Derecho Genetico-Eduardo Ayala TandazoJose Eduardo Ayala TandazoAún no hay calificaciones
- Genes HoxDocumento3 páginasGenes HoxJordan aronAún no hay calificaciones
- annotated-PRACTICA 3Documento10 páginasannotated-PRACTICA 3Andreita Estefany Ventura TejadaAún no hay calificaciones
- Rvou1 OdontogenDocumento3 páginasRvou1 OdontogenkeithgaymerAún no hay calificaciones
- CoronaCruz LIno M16S4PIDocumento4 páginasCoronaCruz LIno M16S4PIDiDi HigieneAún no hay calificaciones
- Desarrollo Neuroembriológico: El Camino Desde La Proliferación Hasta La PerfecciónDocumento11 páginasDesarrollo Neuroembriológico: El Camino Desde La Proliferación Hasta La Perfecciónmaria_daniAún no hay calificaciones
- Evolución Cognitiva en Primates FF 95Documento13 páginasEvolución Cognitiva en Primates FF 95Mateo QuecanoAún no hay calificaciones
- Arbol Filogenetico Bases Biológicas de La Conducta Ana Patricia LBDocumento4 páginasArbol Filogenetico Bases Biológicas de La Conducta Ana Patricia LBAnna BustamanteAún no hay calificaciones
- Actividad de Los Genes Tipo msx-1 Durante El Desarrollo Craneofacial PDFDocumento5 páginasActividad de Los Genes Tipo msx-1 Durante El Desarrollo Craneofacial PDFElizabeth HerhuayAún no hay calificaciones
- The Origin of Metazoa A Unicellular Perspective - En.esDocumento17 páginasThe Origin of Metazoa A Unicellular Perspective - En.esManuel garcia rosasAún no hay calificaciones
- El Reloj OváricoDocumento18 páginasEl Reloj OváricocatatoumarocoAún no hay calificaciones
- Embriologia HipofisisDocumento13 páginasEmbriologia HipofisismiguelalmenarezAún no hay calificaciones
- El Desarrollo Embrionario y Los Genes HomeoticosDocumento3 páginasEl Desarrollo Embrionario y Los Genes HomeoticosLuis Espinoza NAún no hay calificaciones
- Fisiologia AnimalDocumento23 páginasFisiologia AnimalEnrique TapiaAún no hay calificaciones
- b2 Act10Documento2 páginasb2 Act10dntwAún no hay calificaciones
- Ensayo Del Genoma Humano 22-9Documento7 páginasEnsayo Del Genoma Humano 22-9Aura Dilia Flores BaezAún no hay calificaciones
- Dialnet DeLaGeneticaDeLaMoscaALaSaludHumana 2865151Documento7 páginasDialnet DeLaGeneticaDeLaMoscaALaSaludHumana 2865151TabathaAún no hay calificaciones
- FOR - LABIO.SUP. .En - Es PDFDocumento25 páginasFOR - LABIO.SUP. .En - Es PDFYady S. Londoño L.Aún no hay calificaciones
- La FilogeniaDocumento11 páginasLa FilogeniaMishaly PalaciosAún no hay calificaciones
- Progreso en El Conocimiento de La Microbiota Intestinal HumanaDocumento5 páginasProgreso en El Conocimiento de La Microbiota Intestinal HumanaJoel FernandezAún no hay calificaciones
- Resumen Biologia Humana 3Documento61 páginasResumen Biologia Humana 3LucianaAún no hay calificaciones
- Manuel Meruane. Desarrollo de Cara y Cuello en Vertebrados. International Journal of Morphology, 2012 30 (4) 1373-1388.Documento17 páginasManuel Meruane. Desarrollo de Cara y Cuello en Vertebrados. International Journal of Morphology, 2012 30 (4) 1373-1388.Vasquez RAún no hay calificaciones
- TEMA 9 (Psicobiología)Documento5 páginasTEMA 9 (Psicobiología)Natalia González MateosAún no hay calificaciones
- 840 2370 1 PBDocumento18 páginas840 2370 1 PBYo MeAún no hay calificaciones
- Microbioma IntestinalDocumento11 páginasMicrobioma IntestinalAdes GuaremaAún no hay calificaciones
- AcroporaDocumento20 páginasAcroporaYamile A Rodríguez RiascosAún no hay calificaciones
- 54855-Text de L'article-66454-1-10-20070314Documento4 páginas54855-Text de L'article-66454-1-10-20070314Daniela FAún no hay calificaciones
- Consulta 6Documento6 páginasConsulta 6Angy PavónAún no hay calificaciones
- Bioquimica 2Documento9 páginasBioquimica 2mariaisabel_1807Aún no hay calificaciones
- Apunte Embriología IDocumento2 páginasApunte Embriología INatyta GutierrezAún no hay calificaciones
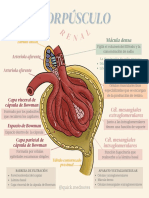
- Corpúsculo Renal - MedicinaDocumento1 páginaCorpúsculo Renal - MedicinaNatyta GutierrezAún no hay calificaciones
- Tablas de Constantes y Otras CosasDocumento4 páginasTablas de Constantes y Otras CosasNatyta GutierrezAún no hay calificaciones
- Nomenclatura en Quimica InorganicaDocumento68 páginasNomenclatura en Quimica InorganicaNatyta GutierrezAún no hay calificaciones
- Equilibrio Acidobásico en El Humano Páginas DesdePratt - Bioquimica 2da Ed 2012Documento2 páginasEquilibrio Acidobásico en El Humano Páginas DesdePratt - Bioquimica 2da Ed 2012Natyta GutierrezAún no hay calificaciones
- Resumen EcologíaDocumento16 páginasResumen EcologíaNatyta GutierrezAún no hay calificaciones
- Lectura Bioenergética PDFDocumento5 páginasLectura Bioenergética PDFJavieralfonso EscamillaAún no hay calificaciones
- Ayudas Qmca OrgánicaDocumento14 páginasAyudas Qmca OrgánicaNatyta GutierrezAún no hay calificaciones
- Resumen EcologíaDocumento16 páginasResumen EcologíaNatyta GutierrezAún no hay calificaciones
- El CarbonoDocumento2 páginasEl CarbonoNatyta GutierrezAún no hay calificaciones
- Unidades de Concentración ResumenDocumento1 páginaUnidades de Concentración ResumenNatyta GutierrezAún no hay calificaciones
- Ayudas Qmca OrgánicaDocumento14 páginasAyudas Qmca OrgánicaNatyta GutierrezAún no hay calificaciones
- El CarbonoDocumento2 páginasEl CarbonoNatyta GutierrezAún no hay calificaciones
- Resumen GasesDocumento1 páginaResumen GasesNatyta GutierrezAún no hay calificaciones
- Tarea en Progreso Completada Cancelada Cumpleaños Evento Cita Importante Pregunta Idea MigradaDocumento1 páginaTarea en Progreso Completada Cancelada Cumpleaños Evento Cita Importante Pregunta Idea MigradaNatyta Gutierrez100% (1)
- Agenda 2020aDocumento1 páginaAgenda 2020aNatyta GutierrezAún no hay calificaciones
- Calendario MensualDocumento13 páginasCalendario MensualNatyta GutierrezAún no hay calificaciones
- Bujo NatyDocumento128 páginasBujo NatyNatyta GutierrezAún no hay calificaciones
- Fases Lunares 2020Documento2 páginasFases Lunares 2020Natyta GutierrezAún no hay calificaciones
- Manual de Procedimientos Tecnicos para El Diagnostico Micologico - FinalDocumento139 páginasManual de Procedimientos Tecnicos para El Diagnostico Micologico - FinalIgnacio Candia Ramos100% (1)
- Informe de Radiosensibilidad - Reyes Ci.28442084Documento4 páginasInforme de Radiosensibilidad - Reyes Ci.28442084JoseAún no hay calificaciones
- EXCRECIONDocumento4 páginasEXCRECIONAmparo OssaAún no hay calificaciones
- Objetivos GeneralesDocumento5 páginasObjetivos GeneralesmartovpAún no hay calificaciones
- BioquimicaDocumento12 páginasBioquimicaVegas PAún no hay calificaciones
- Taller de FarmacologiaDocumento6 páginasTaller de FarmacologiaCesar ArguelloAún no hay calificaciones
- Neuro TrasmisoresDocumento5 páginasNeuro Trasmisorescatalina acosta100% (1)
- Protocolo de La Práctica de Laboratorio de BiologíaDocumento23 páginasProtocolo de La Práctica de Laboratorio de BiologíaJuanAún no hay calificaciones
- U CatolicaDocumento38 páginasU CatolicaCascadasDelCristalAún no hay calificaciones
- Criterios Tecnicos APMES MSPS IDocumento6 páginasCriterios Tecnicos APMES MSPS IMargarita PorrasAún no hay calificaciones
- Camelidos SudamericanosDocumento4 páginasCamelidos SudamericanosMiriam Vilcatoma SotacuroAún no hay calificaciones
- Morgan 3 Unidad 1Documento14 páginasMorgan 3 Unidad 1Luciana CalabreseAún no hay calificaciones
- FisiologíaDocumento28 páginasFisiologíaF.C. BorrásAún no hay calificaciones
- Informe Ing Edson Covid 19Documento5 páginasInforme Ing Edson Covid 19Richard PerezAún no hay calificaciones
- Diferencias Entre Arqueas y Bacterias - DiferenciadorDocumento10 páginasDiferencias Entre Arqueas y Bacterias - DiferenciadorolgaAún no hay calificaciones
- Informe CrustaceosDocumento6 páginasInforme CrustaceosPetete YeYoAún no hay calificaciones
- Epítome Tomo IDocumento268 páginasEpítome Tomo IAlejandro Hernandez PeraltaAún no hay calificaciones
- Biologia - DIMENSION VERDE GRADO 10Documento10 páginasBiologia - DIMENSION VERDE GRADO 10Shellen Baez100% (1)
- Biologia Articulo InformativoDocumento14 páginasBiologia Articulo InformativoDavid Alejandro JimenezAún no hay calificaciones
- Bioquimica Juan CarbohidratosDocumento4 páginasBioquimica Juan CarbohidratosJandry ZambranoAún no hay calificaciones
- BOT FinalDocumento81 páginasBOT FinalFlora MiguelizAún no hay calificaciones
- Inmunidad y Ejercicio PDFDocumento66 páginasInmunidad y Ejercicio PDFOswaldo Quezada TorrezAún no hay calificaciones
- Webinar 01.09.2020Documento27 páginasWebinar 01.09.2020Valeska Andrea Muñoz VelasquezAún no hay calificaciones
- Dimension Biologica de La EnfermedadDocumento24 páginasDimension Biologica de La Enfermedadkcc2850% (2)
- Generalidades HistologíaDocumento16 páginasGeneralidades HistologíaDuvan VargasAún no hay calificaciones
- Jugo GástricoDocumento8 páginasJugo GástricoPriscilla Sandoya BernabéAún no hay calificaciones