También podría gustarte
- Tarea IV de PsicologiaDocumento4 páginasTarea IV de PsicologiaEzequiel Castillo MatosAún no hay calificaciones
- Anatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2De EverandAnatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2Aún no hay calificaciones
- Tarea 4 de La Histoira de La PsicologiaDocumento3 páginasTarea 4 de La Histoira de La PsicologiajoanAún no hay calificaciones
- Neuro Cien CIADocumento17 páginasNeuro Cien CIAemilioAún no hay calificaciones
- NEUROCIENCIASDocumento16 páginasNEUROCIENCIASViry ChristensenAún no hay calificaciones
- Comunicación NeuronalDocumento9 páginasComunicación NeuronalCAROLINA ANDREA MORALES TRIBINOAún no hay calificaciones
- Tarea 4 Historia de La PsicologiaDocumento5 páginasTarea 4 Historia de La PsicologiaignacioAún no hay calificaciones
- NeurocienciaDocumento12 páginasNeurocienciaBS3091Aún no hay calificaciones
- Neuropsicologia ArmandoDocumento51 páginasNeuropsicologia Armandonoe83marAún no hay calificaciones
- Historia de La Psicología Tarea 4Documento3 páginasHistoria de La Psicología Tarea 4Norenny CadetAún no hay calificaciones
- Neurociencias ColombiaDocumento6 páginasNeurociencias ColombiaJuan Pablo CruzAún no hay calificaciones
- Sistemas de Memoria2Documento19 páginasSistemas de Memoria2Eduardo Blanco CalvoAún no hay calificaciones
- Antecedentes de Neuropsicología InfantílDocumento33 páginasAntecedentes de Neuropsicología InfantílNicte RodriguezAún no hay calificaciones
- 1 Parcial NeuroDocumento5 páginas1 Parcial NeuroAgustina PiccoAún no hay calificaciones
- Bases Neuologicas Del ComportamientoDocumento397 páginasBases Neuologicas Del ComportamientoArturo ContrerasAún no hay calificaciones
- Bases Psicobiologicas Del ComportamientoDocumento80 páginasBases Psicobiologicas Del ComportamientoJorge Contreras Garcia IAún no hay calificaciones
- Todo Sobre Memoria y Aprendizaje IDocumento16 páginasTodo Sobre Memoria y Aprendizaje ILilia GarciaAún no hay calificaciones
- Neurona TrabajoDocumento9 páginasNeurona TrabajoMalvina Soledad TorresAún no hay calificaciones
- Tarea 4 Historia de La PsicologiaDocumento4 páginasTarea 4 Historia de La Psicologiameraris chapman100% (4)
- Actividad de La Unidad IVDocumento3 páginasActividad de La Unidad IVJenny Molina de Ignacio67% (3)
- SjskdjiwnshdiskdnsjDocumento24 páginasSjskdjiwnshdiskdnsjWang LinAún no hay calificaciones
- Clase 1 NeurocienciaDocumento16 páginasClase 1 NeurocienciaElectricidad AutomotrizAún no hay calificaciones
- Tema 1Documento14 páginasTema 1nuriazr.08Aún no hay calificaciones
- Fichas 3. Sistema Nervioso, Siapsis y Estructura, Dos Fichas de Parafrasis y Una de ResumenDocumento7 páginasFichas 3. Sistema Nervioso, Siapsis y Estructura, Dos Fichas de Parafrasis y Una de ResumenIrving SanchezAún no hay calificaciones
- Ramón y Cajal y La Neurociencia Del Siglo XXI RESUMENDocumento2 páginasRamón y Cajal y La Neurociencia Del Siglo XXI RESUMENJoel CoriaAún no hay calificaciones
- AntologÍa NeurofisiologÍa IIDocumento37 páginasAntologÍa NeurofisiologÍa IIabrilosegueraAún no hay calificaciones
- Las Redes CorticalesDocumento30 páginasLas Redes CorticalesGiovanni Gabriel100% (4)
- Biología 4to Parcial Proyecto 1Documento3 páginasBiología 4to Parcial Proyecto 1Anthony GuerraAún no hay calificaciones
- El Cerebro y El ComportamientoDocumento17 páginasEl Cerebro y El ComportamientoKatherin castellanosAún no hay calificaciones
- Aporte y Campo de Acción de Las NeurocienciasDocumento11 páginasAporte y Campo de Acción de Las Neurocienciasmarcosola67% (3)
- Tema 5Documento11 páginasTema 5JtavidAún no hay calificaciones
- Fundamentos PsicologicosDocumento14 páginasFundamentos Psicologicoswilmer_1981Aún no hay calificaciones
- El Contexto NeurofisiológicoDocumento4 páginasEl Contexto NeurofisiológicoEDAURYAún no hay calificaciones
- Capitulo 2 Diojani SepulvedaDocumento7 páginasCapitulo 2 Diojani SepulvedaDiojani Sepulveda Terrero100% (1)
- NeurobiologíaDocumento8 páginasNeurobiologíaSylwia AnnaAún no hay calificaciones
- Neuroplasticidad O.foster M.terzaghiDocumento16 páginasNeuroplasticidad O.foster M.terzaghinicole50% (2)
- NeurocienciaDocumento13 páginasNeurocienciaGénesis SangronisAún no hay calificaciones
- Aporte A La NeurocienciaDocumento6 páginasAporte A La Neurocienciaalexis_isaac96Aún no hay calificaciones
- Historia de La Psicologia Unidad IVDocumento4 páginasHistoria de La Psicologia Unidad IVGriselMonegroAún no hay calificaciones
- Apunte 1 UltraestructuraDocumento7 páginasApunte 1 UltraestructuraRafahela Antonia Vergara RiveraAún no hay calificaciones
- Definición y Breve Historia de La NeuropsicologíaDocumento5 páginasDefinición y Breve Historia de La NeuropsicologíaDiana Rodriguez100% (1)
- Tarea 4 Historia de La PsicologiaDocumento4 páginasTarea 4 Historia de La Psicologiaanon_462558415100% (4)
- Unidad 1Documento32 páginasUnidad 1ymarozziAún no hay calificaciones
- NEURO1Documento26 páginasNEURO1tonyx100% (1)
- Cap 1 - Elementos Propedéuticos de Neuropsicologia PDFDocumento19 páginasCap 1 - Elementos Propedéuticos de Neuropsicologia PDFKILAMAZUKEAún no hay calificaciones
- Las Neuronas - Fundamentos y Funciones - Nayomi YacorDocumento2 páginasLas Neuronas - Fundamentos y Funciones - Nayomi YacorNayomiAún no hay calificaciones
- La Quimica Del Cerebro Ccesa07Documento135 páginasLa Quimica Del Cerebro Ccesa07Demetrio Ccesa RaymeAún no hay calificaciones
- La Química Del Pensamiento: Dr. Fernando CabiesesDocumento3 páginasLa Química Del Pensamiento: Dr. Fernando Cabiesesonurb90Aún no hay calificaciones
- Resumen Elementos de La Neuropsicologia 2021Documento81 páginasResumen Elementos de La Neuropsicologia 2021Alejandra Feliz DidierAún no hay calificaciones
- Neurociencia 4Documento38 páginasNeurociencia 4Luis CahuanaAún no hay calificaciones
- Reporte de Las 3 LecturasDocumento23 páginasReporte de Las 3 Lecturasnallely vidalAún no hay calificaciones
- Resumen NeurocienciasDocumento9 páginasResumen NeurocienciasbriAún no hay calificaciones
- Origen de La NeurocienciaDocumento7 páginasOrigen de La NeurocienciaVictor Díaz AlarconAún no hay calificaciones
- Aprendizaje y MemoriaDocumento14 páginasAprendizaje y MemoriaElfroguerAún no hay calificaciones
- Sensación Y PercepciónDocumento8 páginasSensación Y PercepciónMilagros CarvajalAún no hay calificaciones
- 1°parcial ResumenDocumento20 páginas1°parcial Resumenminnie.kim1807Aún no hay calificaciones
- Tema 1 Apuntes Hechos DefinitivosDocumento17 páginasTema 1 Apuntes Hechos Definitivosbertin osborneAún no hay calificaciones
- Ontogenia de La Auto Con Ciencia. Como Se Contruye El Cerebro CognitivoDocumento6 páginasOntogenia de La Auto Con Ciencia. Como Se Contruye El Cerebro CognitivoMefisto Dolores DesaucioAún no hay calificaciones
- Proyecto Oso Panda Aula 3 Ac3b1osDocumento19 páginasProyecto Oso Panda Aula 3 Ac3b1osmonica gAún no hay calificaciones
- Revista Etologia Del FelinoDocumento16 páginasRevista Etologia Del FelinoPaula Galimidi Valenzuela0% (1)
- Ferreira NeurocienciaDocumento94 páginasFerreira NeurocienciaJorge MartinezAún no hay calificaciones
- El Distrés y El EustrésDocumento2 páginasEl Distrés y El EustréssandraAún no hay calificaciones
- Taller de Genetica Mendelian1Documento15 páginasTaller de Genetica Mendelian1Alejandra Clavijo AristizabalAún no hay calificaciones
- Informacion Seminario Tejido NerviosoDocumento7 páginasInformacion Seminario Tejido NerviosoRuben PeñaAún no hay calificaciones
- Iv. Materiales Y Metodos 4.1. MaterialesDocumento10 páginasIv. Materiales Y Metodos 4.1. MaterialeslyfaAún no hay calificaciones
- ESTO ATERRA - Jose Luis TaverasDocumento2 páginasESTO ATERRA - Jose Luis Taveraspharah vidalAún no hay calificaciones
- Regulacion GenicaDocumento2 páginasRegulacion Genicajuan david perez posadasAún no hay calificaciones
- Contaminacion AmbientalDocumento4 páginasContaminacion AmbientalStefani Cabrera PortocarreroAún no hay calificaciones
- Vision Presentacion FinalDocumento27 páginasVision Presentacion FinalYordano CRAún no hay calificaciones
- Los 5 Reinos de La NaturalezaDocumento1 páginaLos 5 Reinos de La NaturalezaLourdes BrionesAún no hay calificaciones
- 7 DermatofitosDocumento107 páginas7 DermatofitosRosmeri Melani100% (1)
- Aparato ReproductorDocumento8 páginasAparato ReproductorDavid Ignacio Reyes MurciaAún no hay calificaciones
- Fisiologia Vegetal - 30 NoviembreDocumento3 páginasFisiologia Vegetal - 30 NoviembreOscar Alarcon JimenezAún no hay calificaciones
- Trabajo QuimicaDocumento16 páginasTrabajo QuimicaDavid GuidoAún no hay calificaciones
- Actividades Complementarias BiologíaDocumento89 páginasActividades Complementarias BiologíaYorlan Cervantes PerezAún no hay calificaciones
- Cura para El HONGO UÑADocumento5 páginasCura para El HONGO UÑANelly BeatrizAún no hay calificaciones
- Trabajo Práctico N 3Documento4 páginasTrabajo Práctico N 3Celina ContrerasAún no hay calificaciones
- Tabla de Eones Periodos y EpocasDocumento3 páginasTabla de Eones Periodos y EpocasDoris Marlene MolinaAún no hay calificaciones
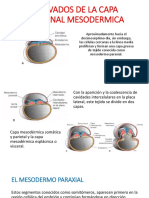
- Derivados de La Capa Germinal MesodermicaDocumento15 páginasDerivados de La Capa Germinal Mesodermicayanethherrera0% (1)
- Concepto de Compartimientos LeucocitariosDocumento3 páginasConcepto de Compartimientos LeucocitariosCarlos Lopez Gonzales100% (2)
- Repaso EspanolDocumento7 páginasRepaso EspanolAndrea100% (1)
- Aparato Reproductor MasculinoDocumento1 páginaAparato Reproductor MasculinoArturo AhumadaAún no hay calificaciones
- Poteciales Evocados - Tema 7Documento22 páginasPoteciales Evocados - Tema 7María Fernanda LamusAún no hay calificaciones
- La CodominanciaDocumento3 páginasLa CodominanciaSeries droidAún no hay calificaciones
- Sistema LinfáticoDocumento18 páginasSistema LinfáticoIslendy Paola Álvarez LondoñoAún no hay calificaciones
- AsfixiaDocumento50 páginasAsfixiaDivisay VenenciaAún no hay calificaciones
- Manual de Captación y Aprovechamiento Del Agua de Lluvia - FAODocumento235 páginasManual de Captación y Aprovechamiento Del Agua de Lluvia - FAOpiquiriderAún no hay calificaciones
- Ficha Tecnica RutinalDocumento1 páginaFicha Tecnica Rutinalandres vargasAún no hay calificaciones
- Ejercicios de Psicología Positiva para aumentar tu felicidadDe EverandEjercicios de Psicología Positiva para aumentar tu felicidadCalificación: 4.5 de 5 estrellas4.5/5 (18)
- Homo antecessor: El nacimiento de una especieDe EverandHomo antecessor: El nacimiento de una especieCalificación: 5 de 5 estrellas5/5 (1)
- Entrevista clínica y psicometríaDe EverandEntrevista clínica y psicometríaCalificación: 4.5 de 5 estrellas4.5/5 (5)
- Manual de psicoterapia emocional sistémicaDe EverandManual de psicoterapia emocional sistémicaCalificación: 4.5 de 5 estrellas4.5/5 (7)
- El poder de la presencia: Cómo la presencia de los padres moldea el cerebro de los hijos y configura las personas que llegarán a serDe EverandEl poder de la presencia: Cómo la presencia de los padres moldea el cerebro de los hijos y configura las personas que llegarán a serCalificación: 4.5 de 5 estrellas4.5/5 (11)
- Introducción a la psicología generalDe EverandIntroducción a la psicología generalCalificación: 3.5 de 5 estrellas3.5/5 (16)
- Educar la atención: Cómo entrenar esta habilidad en niños y adultosDe EverandEducar la atención: Cómo entrenar esta habilidad en niños y adultosCalificación: 4.5 de 5 estrellas4.5/5 (10)
- Cuerpo Tóxico: Como Liberar Tu Cuerpo De Las Toxinas Externas E Internas, Y Evitar Asi Los Efectos De Los Radicales LibresDe EverandCuerpo Tóxico: Como Liberar Tu Cuerpo De Las Toxinas Externas E Internas, Y Evitar Asi Los Efectos De Los Radicales LibresCalificación: 5 de 5 estrellas5/5 (2)
- Inteligencia artificial: Lo que usted necesita saber sobre el aprendizaje automático, robótica, aprendizaje profundo, Internet de las cosas, redes neuronales, y nuestro futuroDe EverandInteligencia artificial: Lo que usted necesita saber sobre el aprendizaje automático, robótica, aprendizaje profundo, Internet de las cosas, redes neuronales, y nuestro futuroCalificación: 4 de 5 estrellas4/5 (1)
- 50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaDe Everand50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaCalificación: 4 de 5 estrellas4/5 (37)
- Anatomía del caballo: Guía práctica ilustradaDe EverandAnatomía del caballo: Guía práctica ilustradaCalificación: 4 de 5 estrellas4/5 (9)
- Mapas Mentales ¡La Guía del Principiante Para Crear Mapas Mentales Paso a Paso!De EverandMapas Mentales ¡La Guía del Principiante Para Crear Mapas Mentales Paso a Paso!Calificación: 3.5 de 5 estrellas3.5/5 (16)
- Sesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesDe EverandSesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesCalificación: 4.5 de 5 estrellas4.5/5 (13)
- Liberación del trauma: Perdón y temblor es el caminoDe EverandLiberación del trauma: Perdón y temblor es el caminoCalificación: 4 de 5 estrellas4/5 (4)
- Cómo conversar con cualquiera: Descubre cómo ser la persona más interesante del lugar y a destruir la ansiedad social permanentementeDe EverandCómo conversar con cualquiera: Descubre cómo ser la persona más interesante del lugar y a destruir la ansiedad social permanentementeCalificación: 4 de 5 estrellas4/5 (81)
- Ansiedad: Vencer La Ansiedad, El Miedo, El Estrés Y La Depresión, Y Deshacerse De Las FobiasDe EverandAnsiedad: Vencer La Ansiedad, El Miedo, El Estrés Y La Depresión, Y Deshacerse De Las FobiasCalificación: 4.5 de 5 estrellas4.5/5 (17)
- Las Enfermedades comienzan y terminan en tu mente: Una guía para la autosanaciónDe EverandLas Enfermedades comienzan y terminan en tu mente: Una guía para la autosanaciónCalificación: 4 de 5 estrellas4/5 (4)
- Autoestima: Cómo evaluar, mejorar y mantener una sana autoestimaDe EverandAutoestima: Cómo evaluar, mejorar y mantener una sana autoestimaCalificación: 4.5 de 5 estrellas4.5/5 (10)
- Trabajo con padres y terapia con hijos: Un modelo integradorDe EverandTrabajo con padres y terapia con hijos: Un modelo integradorCalificación: 5 de 5 estrellas5/5 (2)
- Apego y Terapia Narrativa: un modelo integradorDe EverandApego y Terapia Narrativa: un modelo integradorCalificación: 4.5 de 5 estrellas4.5/5 (6)
- Cerebro y silencio: Las claves de la creatividad y la serenidadDe EverandCerebro y silencio: Las claves de la creatividad y la serenidadCalificación: 5 de 5 estrellas5/5 (2)
- Resumen de Pensar rápido pensar despacio de Daniel KahnemanDe EverandResumen de Pensar rápido pensar despacio de Daniel KahnemanCalificación: 4.5 de 5 estrellas4.5/5 (11)
- Parásitos: El extraño mundo de las criaturas más peligrosas de la naturalezaDe EverandParásitos: El extraño mundo de las criaturas más peligrosas de la naturalezaCalificación: 4.5 de 5 estrellas4.5/5 (47)
- La conciencia en el cerebro: Descifrando el enigma de cómo el cerebro elabora nuestros pensamientosDe EverandLa conciencia en el cerebro: Descifrando el enigma de cómo el cerebro elabora nuestros pensamientosCalificación: 2.5 de 5 estrellas2.5/5 (3)