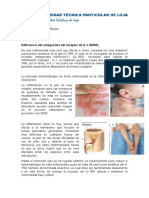
También podría gustarte
- Identificación de hongos asociados a suelos agrícolasDe EverandIdentificación de hongos asociados a suelos agrícolasAún no hay calificaciones
- Manejo agroecológico de la mosca blanca en el cultivo de berenjenaDe EverandManejo agroecológico de la mosca blanca en el cultivo de berenjenaCalificación: 5 de 5 estrellas5/5 (1)
- Álvarez López 2013Documento14 páginasÁlvarez López 2013Francisco CárdenasAún no hay calificaciones
- Establecimiento in Vitro de Segmentos Nodales de Guanabana 2020Documento7 páginasEstablecimiento in Vitro de Segmentos Nodales de Guanabana 2020ANGELA DANIELA CANSECO TARIFAAún no hay calificaciones
- Informe de Laboratorio 7Documento10 páginasInforme de Laboratorio 7Yeinis EscorciaAún no hay calificaciones
- Potencial Biofertilizante de Bacterias Aisladada Del Cultivo Del CacaoDocumento9 páginasPotencial Biofertilizante de Bacterias Aisladada Del Cultivo Del CacaoBRAYAN FELIPE GONZALES QUIGUANASAún no hay calificaciones
- Propuesta Trabajo Grado Michael Final Revisado Elkin JimenaDocumento4 páginasPropuesta Trabajo Grado Michael Final Revisado Elkin JimenaStiven G. AlvarezAún no hay calificaciones
- Aislamiento e identificación de 10 cepas desnitrificantesDocumento5 páginasAislamiento e identificación de 10 cepas desnitrificantesMauricio Rjas55% (40)
- ARTICULO CIENTIFICO FINCyTDocumento11 páginasARTICULO CIENTIFICO FINCyTStephany Mariela BermudezAún no hay calificaciones
- ArticuloDocumento8 páginasArticuloMAX ALEXANDER NAVARRO CHUQUIYAURIAún no hay calificaciones
- 889-Texto Del Artículo-2773-2-10-20231222Documento6 páginas889-Texto Del Artículo-2773-2-10-20231222ALVARO MAURICIO CEVALLOS RAMIREZAún no hay calificaciones
- NR41777 en EsDocumento10 páginasNR41777 en EsLaura FerrerAún no hay calificaciones
- 647-Texto Del Artículo-1661-1-10-20210714Documento4 páginas647-Texto Del Artículo-1661-1-10-20210714Yenny nyceAún no hay calificaciones
- 647-Texto Del Artículo-1661-1-10-20210714Documento4 páginas647-Texto Del Artículo-1661-1-10-20210714Yenny nyceAún no hay calificaciones
- Articulo+18_2130Documento10 páginasArticulo+18_2130teresita.arizaAún no hay calificaciones
- DESARROLLO DE PLANTAS DE MOSTAZA (Brassica Juncea) CULTIVADAS EN SUELOS y JALES MINEROSDocumento6 páginasDESARROLLO DE PLANTAS DE MOSTAZA (Brassica Juncea) CULTIVADAS EN SUELOS y JALES MINEROSLuis CastroAún no hay calificaciones
- Aislamiento y Evaluación de Bacterias (Grupo N°1)Documento5 páginasAislamiento y Evaluación de Bacterias (Grupo N°1)Rosalinda Apaza apazaAún no hay calificaciones
- Tarea 2 QuishpeDocumento31 páginasTarea 2 Quishpemayerli rashel garcia vilañaAún no hay calificaciones
- Informe - Suspension Celular ZanahoriaDocumento4 páginasInforme - Suspension Celular ZanahoriaPatricia TorresAún no hay calificaciones
- Seleccion de Bacterias Fluorescentes ProductorasDocumento13 páginasSeleccion de Bacterias Fluorescentes Productorasviwef68893Aún no hay calificaciones
- Caracterización morfológica y molecular de la colección nacional de camote en EcuadorDocumento5 páginasCaracterización morfológica y molecular de la colección nacional de camote en EcuadorRichard Molina AlvarezAún no hay calificaciones
- Efecto de Reguladores de Crecimiento en PlantasDocumento15 páginasEfecto de Reguladores de Crecimiento en PlantasCarolina CarbajalAún no hay calificaciones
- Casos de BiorremediaciónDocumento15 páginasCasos de Biorremediaciónjohana paola martinez albarracinAún no hay calificaciones
- Evaluacion Microbiologica de Un Policultivo de Lombrices de Tierra en La Depuracion Del Residual PorcinoDocumento3 páginasEvaluacion Microbiologica de Un Policultivo de Lombrices de Tierra en La Depuracion Del Residual PorcinoFabian Ignacio Abarza VillalobosAún no hay calificaciones
- Trabajo II de Osbel Núñez, Atagua (1) - 0410113159Documento11 páginasTrabajo II de Osbel Núñez, Atagua (1) - 0410113159Joyssi LopezAún no hay calificaciones
- Procedimiento Experimental para La Determinación de Bacterias Solubilizadoras de Fosfato DESTACADODocumento5 páginasProcedimiento Experimental para La Determinación de Bacterias Solubilizadoras de Fosfato DESTACADOSebastian Armijo MendozaAún no hay calificaciones
- 1819 4087 CTR 41 02 E03Documento17 páginas1819 4087 CTR 41 02 E03Dayana Arce LopezAún no hay calificaciones
- Paso 3 Obtener La Información de Las Estrategias de ManejoDocumento11 páginasPaso 3 Obtener La Información de Las Estrategias de Manejoluis barretoAún no hay calificaciones
- CultivoDocumento7 páginasCultivoCarolina Panchana TAún no hay calificaciones
- Introduccion y Multiplicacion in Vitro de Morado BarranquinoDocumento4 páginasIntroduccion y Multiplicacion in Vitro de Morado BarranquinoMilagros Del Rosario Palacios VellonAún no hay calificaciones
- Camacho 2015Documento15 páginasCamacho 2015Francisco CárdenasAún no hay calificaciones
- Verificación y CuantificaciónDocumento4 páginasVerificación y CuantificaciónPq Adolfo MichaelAún no hay calificaciones
- Plantilla BannerDocumento15 páginasPlantilla BannerCristhian Coila DiazAún no hay calificaciones
- Aislamiento de Bacterias NitrificantesDocumento4 páginasAislamiento de Bacterias Nitrificantesbr.mateomateoAún no hay calificaciones
- MT Establecimiento y Multiplicación in Vitro de Embriones GuanabanaDocumento10 páginasMT Establecimiento y Multiplicación in Vitro de Embriones GuanabanaPuro HuesoAún no hay calificaciones
- Anteproyecto de Tesis - Marchan Zapata MarcioDocumento22 páginasAnteproyecto de Tesis - Marchan Zapata MarcioMarcio MarchanAún no hay calificaciones
- CONICET Digital Nro. A PDFDocumento256 páginasCONICET Digital Nro. A PDFPepoAún no hay calificaciones
- 1819-4087-ctr-40-02-e05Documento29 páginas1819-4087-ctr-40-02-e05Andreina ZamoraAún no hay calificaciones
- Cultivo de meristemos apicalesDocumento8 páginasCultivo de meristemos apicalesPatricia TorresAún no hay calificaciones
- Informe de Laboratorio 4-FBN SimbioticasDocumento7 páginasInforme de Laboratorio 4-FBN SimbioticasjeancAún no hay calificaciones
- FITORREMEDIACIONDocumento58 páginasFITORREMEDIACIONXharitto TCAún no hay calificaciones
- Informe #2Documento13 páginasInforme #2nohoraAún no hay calificaciones
- Propagación clonal in vitro de tecaDocumento6 páginasPropagación clonal in vitro de tecaElla OdelantiAún no hay calificaciones
- SAEZ 2023 - Resumen PIN II 04 A147Documento1 páginaSAEZ 2023 - Resumen PIN II 04 A147Julieta SaezAún no hay calificaciones
- Rhizopua Stolonifer. Efecto en Los Diferentes Medios de Cultivo en La Produccion de Biomasa y Esgosterol.Documento7 páginasRhizopua Stolonifer. Efecto en Los Diferentes Medios de Cultivo en La Produccion de Biomasa y Esgosterol.Ed, Edd y Eddy HernándezAún no hay calificaciones
- Microorganismos Beneficos Como Biofertilizantes Eficientes para El Cultivo Del TomateDocumento8 páginasMicroorganismos Beneficos Como Biofertilizantes Eficientes para El Cultivo Del TomateLuis EduardoAún no hay calificaciones
- AZOTOBACTERDocumento3 páginasAZOTOBACTEREstefania MacorAún no hay calificaciones
- Efecto del biocarbón de plátano en la disponibilidad de arsénico en arrozDocumento22 páginasEfecto del biocarbón de plátano en la disponibilidad de arsénico en arrozMarcio MarchanAún no hay calificaciones
- Investigación de Metabolitos Secundarios Producidos Por Hongos Filamentosos Recuperados de Tucumán y Su Potencial en La IndustriaDocumento8 páginasInvestigación de Metabolitos Secundarios Producidos Por Hongos Filamentosos Recuperados de Tucumán y Su Potencial en La IndustriaTomas5102Aún no hay calificaciones
- Guia de Laboratorio Aislamiento e Identificacion de Clostridium SPPDocumento5 páginasGuia de Laboratorio Aislamiento e Identificacion de Clostridium SPPLEIDE VANESSA GONZALEZ RUEDAAún no hay calificaciones
- Bacterias celulolíticas aisladas del intestino de termitas con potencial probiótico y degradación de pastoDocumento15 páginasBacterias celulolíticas aisladas del intestino de termitas con potencial probiótico y degradación de pastoPiero PaoloAún no hay calificaciones
- 1 Aislamiento y Caracterización de RizobiosDocumento10 páginas1 Aislamiento y Caracterización de RizobiosAugusto SanchezAún no hay calificaciones
- Hongos Del Suelo Saprófitos y Patógenos de Plantas ExposicionDocumento48 páginasHongos Del Suelo Saprófitos y Patógenos de Plantas ExposicioncarolinaAún no hay calificaciones
- METODOLOGIADocumento13 páginasMETODOLOGIAMITCHELLE ORTEGONAún no hay calificaciones
- Aislamiento e Identificación de Micorrizas ArbuscularesDocumento6 páginasAislamiento e Identificación de Micorrizas ArbuscularesFabian GomezAún no hay calificaciones
- Proyecto TerpenosDocumento3 páginasProyecto Terpenosmauricio daniel zamora roblesAún no hay calificaciones
- HERNANDEZ J - Articulo Cientifico - Bioprospección de Actinomicetes Productores de Enzimas Hidrolíticas Procedentes de La Zona NorteDocumento14 páginasHERNANDEZ J - Articulo Cientifico - Bioprospección de Actinomicetes Productores de Enzimas Hidrolíticas Procedentes de La Zona NorteKathyBritoAún no hay calificaciones
- Laboratorio 6 Suelos MicorrizadosDocumento9 páginasLaboratorio 6 Suelos Micorrizadosnestorrojas07Aún no hay calificaciones
- Micorriza PiñaDocumento9 páginasMicorriza PiñaStephanie MorrisAún no hay calificaciones
- Revista de Simulacion y Laboratorio V3 N8 1Documento9 páginasRevista de Simulacion y Laboratorio V3 N8 1Paulina H.Aún no hay calificaciones
- 11 MicorrizasDocumento7 páginas11 MicorrizasDiana Estefania Dominguez RiosAún no hay calificaciones
- Determinación de Alcohol Etilico en SangreDocumento2 páginasDeterminación de Alcohol Etilico en SangreBetty Acaro ReyesAún no hay calificaciones
- Usp 37 NF 32 en Espanol Volumen 4 5817 6919pdf 4 PDF FreeDocumento1218 páginasUsp 37 NF 32 en Espanol Volumen 4 5817 6919pdf 4 PDF FreeBetty AcaroAún no hay calificaciones
- Feum 11 Tomo IiDocumento1493 páginasFeum 11 Tomo IiJavier Galvan92% (84)
- Farmacopea Argentina 2013 Ed.7Documento2745 páginasFarmacopea Argentina 2013 Ed.7Doménica Palacios100% (1)
- Enfermedades InmunológicasDocumento3 páginasEnfermedades InmunológicasBetty AcaroAún no hay calificaciones
- Efecto de La Aplicación de Roca Fosfórica BacteriasDocumento13 páginasEfecto de La Aplicación de Roca Fosfórica BacteriasBetty AcaroAún no hay calificaciones
- UNICEF 2019 - NutricionDocumento8 páginasUNICEF 2019 - NutricionBetty AcaroAún no hay calificaciones
- CriopreservacionDocumento1 páginaCriopreservacionBetty AcaroAún no hay calificaciones
- Las Células de Las Plantas y Los Hongos Tienen Una Pared Celular Rígida Que Les Protege y Da SoporteDocumento7 páginasLas Células de Las Plantas y Los Hongos Tienen Una Pared Celular Rígida Que Les Protege y Da SoporteBetty AcaroAún no hay calificaciones
- Fuentes de ContaminaciónDocumento1 páginaFuentes de ContaminaciónBetty AcaroAún no hay calificaciones
- ToxicologíaDocumento2 páginasToxicologíaBetty AcaroAún no hay calificaciones
- Ventajas de Los Cultivos CelularesDocumento14 páginasVentajas de Los Cultivos CelularesDani OrtegaAún no hay calificaciones
- Tema 1.COMUNICACIÓN CELULAR Primera PateDocumento2 páginasTema 1.COMUNICACIÓN CELULAR Primera PateBetty AcaroAún no hay calificaciones
- Estructura y funciones del citoesqueletoDocumento2 páginasEstructura y funciones del citoesqueletoBetty AcaroAún no hay calificaciones
- Fisiologia Cap 8 Nivel 2Documento5 páginasFisiologia Cap 8 Nivel 2Betty AcaroAún no hay calificaciones
- Pruebas de Laboratorio Clínico Complementarias para COVID19Documento4 páginasPruebas de Laboratorio Clínico Complementarias para COVID19Betty AcaroAún no hay calificaciones
- Capitulo 20 Fisologia Nivel 2Documento9 páginasCapitulo 20 Fisologia Nivel 2Betty AcaroAún no hay calificaciones
- Infecciones GastrointestinalesDocumento3 páginasInfecciones GastrointestinalesBetty AcaroAún no hay calificaciones
- TOXICOCINETICA CocainaDocumento3 páginasTOXICOCINETICA CocainaBetty AcaroAún no hay calificaciones
- Hemograma: Rangos Normales Glóbulos, Hemoglobina, PlaquetasDocumento5 páginasHemograma: Rangos Normales Glóbulos, Hemoglobina, PlaquetasBetty AcaroAún no hay calificaciones
- ALCOHOLEMIADocumento3 páginasALCOHOLEMIABetty AcaroAún no hay calificaciones
- Capitulo 20 Fisologia Nivel 2Documento9 páginasCapitulo 20 Fisologia Nivel 2Betty AcaroAún no hay calificaciones
- VIRUSDocumento7 páginasVIRUSBetty AcaroAún no hay calificaciones
- Cinchona OfficinalisDocumento2 páginasCinchona OfficinalisBetty AcaroAún no hay calificaciones
- Cinchona OfficinalisDocumento2 páginasCinchona OfficinalisBetty AcaroAún no hay calificaciones
- Resistencia Bacteriana (Bailey & Scott, 2009), y Antibiograma (Velasco, 2008) PDFDocumento21 páginasResistencia Bacteriana (Bailey & Scott, 2009), y Antibiograma (Velasco, 2008) PDFBetty AcaroAún no hay calificaciones
- Resistencia Bacteriana (Bailey & Scott, 2009), y Antibiograma (Velasco, 2008) PDFDocumento21 páginasResistencia Bacteriana (Bailey & Scott, 2009), y Antibiograma (Velasco, 2008) PDFBetty AcaroAún no hay calificaciones
- Movimiento Rectilineo PDFDocumento4 páginasMovimiento Rectilineo PDFBetty AcaroAún no hay calificaciones
- Mapa Conceptual BacteriasDocumento3 páginasMapa Conceptual BacteriasOSCAR FABIAN CANALES GARCIAAún no hay calificaciones
- Mapa Mental MutantesDocumento1 páginaMapa Mental MutantesEstephany BarrigaAún no hay calificaciones
- GenHumExpArgDocumento16 páginasGenHumExpArgJulia Beatriz Alcántara CastilloAún no hay calificaciones
- Actividad 1Documento1 páginaActividad 1Denisse Lizeth Santiago CasaosAún no hay calificaciones
- Microalgas DinoflageladosDocumento23 páginasMicroalgas DinoflageladosJean FrancoAún no hay calificaciones
- Trabajo Practico MutacionesDocumento3 páginasTrabajo Practico Mutacionesbian rotelaAún no hay calificaciones
- Genetica Medica Guias y Clases PracticasDocumento47 páginasGenetica Medica Guias y Clases PracticasAlejandro Malcom Fernández QuintanaAún no hay calificaciones
- Fernandez Vecilla DomingoDocumento34 páginasFernandez Vecilla DomingoMyrian RobertiAún no hay calificaciones
- Informe - Biologia Celular Práctica 10Documento7 páginasInforme - Biologia Celular Práctica 10m12 CAún no hay calificaciones
- Antes de Lucy El Agujero Negro de La Evolucion Humana - AA VVDocumento245 páginasAntes de Lucy El Agujero Negro de La Evolucion Humana - AA VVManuel Benitez100% (1)
- Ensayo Modelo de Sintesis de DNA (ADN)Documento18 páginasEnsayo Modelo de Sintesis de DNA (ADN)Guillermo Enrique Chiunti GarcíaAún no hay calificaciones
- Evaluacion Final C.N.C SolaDocumento6 páginasEvaluacion Final C.N.C SolaHenry Orlando Blanco CarreñoAún no hay calificaciones
- Reproducción Sexual y Asexual Situacion Didactica IDocumento6 páginasReproducción Sexual y Asexual Situacion Didactica IGiuseppe Edward Hernandez Ramos PeraltaAún no hay calificaciones
- Ejercicios - RT-QPCRDocumento12 páginasEjercicios - RT-QPCRKarla ChávezAún no hay calificaciones
- Tipos de ARNDocumento5 páginasTipos de ARNcesar molina canalesAún no hay calificaciones
- Celula VegetalDocumento4 páginasCelula VegetalAlex Ramirez1Aún no hay calificaciones
- Semana 11 CT QuintoDocumento3 páginasSemana 11 CT Quintolaura cerònAún no hay calificaciones
- MACHETE (La Célula Como Unidad de Los Seres Vivos)Documento28 páginasMACHETE (La Célula Como Unidad de Los Seres Vivos)Emilio BernardinoAún no hay calificaciones
- Preguntas Guía Sobre El Texto de MadiganDocumento7 páginasPreguntas Guía Sobre El Texto de MadiganValenthinaCamargoAún no hay calificaciones
- Metabolismo Del ARNDocumento20 páginasMetabolismo Del ARNDavid AnlaAún no hay calificaciones
- Psicologia HumanaDocumento58 páginasPsicologia Humanacarlos peñaAún no hay calificaciones
- Etapas de La Profase I de La Meiosis IDocumento1 páginaEtapas de La Profase I de La Meiosis IJhairo TerronesAún no hay calificaciones
- Especiacion GeranoaetusDocumento7 páginasEspeciacion GeranoaetusJared RicauteAún no hay calificaciones
- Tipos de ReproducciónDocumento2 páginasTipos de ReproducciónFLORES PALESTINA ALEXANDRA0% (1)
- 2°m Taxo 2Documento7 páginas2°m Taxo 2Jessica Lorena DíazAún no hay calificaciones
- Meiosis Mitosis GametogenesisDocumento3 páginasMeiosis Mitosis GametogenesisKEVIN JOSUE NIETO PADILLAAún no hay calificaciones
- Genética Mendeliana (Marielys-Trabajo)Documento18 páginasGenética Mendeliana (Marielys-Trabajo)Jesus RodríguezAún no hay calificaciones
- EVOLUCIÓNDocumento15 páginasEVOLUCIÓNCesar Eneque ChancafeAún no hay calificaciones
- Telomeros y TelomerasasDocumento4 páginasTelomeros y TelomerasasLuz Vanessa ParejoAún no hay calificaciones
- Biologia Informe 5Documento9 páginasBiologia Informe 5salvatoreAún no hay calificaciones