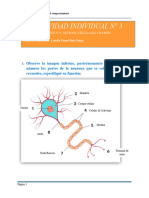
También podría gustarte
- Anatomía y Fisiología del Sistema NerviosoDe EverandAnatomía y Fisiología del Sistema NerviosoCalificación: 4 de 5 estrellas4/5 (19)
- TALLER No. 2 MORFOFISIOLOGÍA - 2 - 2020Documento7 páginasTALLER No. 2 MORFOFISIOLOGÍA - 2 - 2020johanna ramiirezAún no hay calificaciones
- Tejido NerviosoDocumento17 páginasTejido NerviosoMelany MolinaAún no hay calificaciones
- Tarea 3.0 Unidad 3. Control NeuralDocumento25 páginasTarea 3.0 Unidad 3. Control NeuralJuanalkis RodriguezAún no hay calificaciones
- Apunte 4Documento14 páginasApunte 4Ezequiel MAún no hay calificaciones
- Guia Sistema Nervioso 2Documento15 páginasGuia Sistema Nervioso 2FernandoGongora100% (2)
- Guia Sist Nervioso Ciclo 4Documento9 páginasGuia Sist Nervioso Ciclo 4FabianAún no hay calificaciones
- DendritaDocumento6 páginasDendritadsamayoae1Aún no hay calificaciones
- Plasticidad Neural-2003 PDFDocumento9 páginasPlasticidad Neural-2003 PDFAlexis BadillaAún no hay calificaciones
- Guia Sistema Nervioso. SantillanaDocumento13 páginasGuia Sistema Nervioso. SantillanaPedro Nelgui Vargas Quintero100% (2)
- Sistema Nervioso Parte 1Documento42 páginasSistema Nervioso Parte 1Fernando Gabriel Arce TionAún no hay calificaciones
- Tejido NerviosoDocumento52 páginasTejido NerviosoAndrea BaldiviezoAún no hay calificaciones
- FISIOLOGIA - Resumen 1 FisiologíaDocumento68 páginasFISIOLOGIA - Resumen 1 FisiologíaLaura :)Aún no hay calificaciones
- Anatomia y Fisiología Semana 6 - 2021 IiiDocumento8 páginasAnatomia y Fisiología Semana 6 - 2021 IiiDavid LlontopAún no hay calificaciones
- Plasticidad NeuralDocumento0 páginasPlasticidad NeuralSonia FztsAún no hay calificaciones
- 1 Plasticidad NeuralDocumento9 páginas1 Plasticidad NeuralAlejandra LópezAún no hay calificaciones
- Introducción A La Neurociencia - BrainwareDocumento51 páginasIntroducción A La Neurociencia - Brainwaremateo pianaAún no hay calificaciones
- Presentacion Cuerpo Humano Organico Ilustrado Morado PastelDocumento43 páginasPresentacion Cuerpo Humano Organico Ilustrado Morado PastelValeria Gómez0% (1)
- Guia Sistema Nervioso - Juan A SanchezDocumento12 páginasGuia Sistema Nervioso - Juan A SanchezEvelin FontalvoAún no hay calificaciones
- Sistema Nervioso WordDocumento20 páginasSistema Nervioso WordRdioAún no hay calificaciones
- NEUROGLIADocumento16 páginasNEUROGLIADavidAún no hay calificaciones
- Funciones de La Celulas Gliales IDocumento8 páginasFunciones de La Celulas Gliales IJenifer Carolina Pinzón FloresAún no hay calificaciones
- Guia Sistema Nervioso. SantillanaDocumento12 páginasGuia Sistema Nervioso. SantillanaPedro Nelgui Vargas QuinteroAún no hay calificaciones
- Guia Del Sistema Nervioso OctavoDocumento12 páginasGuia Del Sistema Nervioso OctavoRONALD QUIROZAún no hay calificaciones
- Unidad IiDocumento11 páginasUnidad IiCARLAAún no hay calificaciones
- Celulas de Sosten GliaDocumento4 páginasCelulas de Sosten Gliaemil obeidAún no hay calificaciones
- Parcial PsicofisiologíaDocumento10 páginasParcial PsicofisiologíaLeonardo VediaAún no hay calificaciones
- NeurodesarrolloDocumento13 páginasNeurodesarrolloEdmy NavarroAún no hay calificaciones
- Neuronas y Celulas Gliales FichasDocumento5 páginasNeuronas y Celulas Gliales FichasJaviera EricesAún no hay calificaciones
- Anatomia de La NeuronaDocumento3 páginasAnatomia de La NeuronaMoises del CidAún no hay calificaciones
- Transmición SinápticaDocumento2 páginasTransmición Sinápticaalbamarcela0929Aún no hay calificaciones
- Resumen OATec - NeurocienciasDocumento31 páginasResumen OATec - NeurocienciasLucia LlordenAún no hay calificaciones
- FBE Resumen 2Documento6 páginasFBE Resumen 2Fatima Aguilar ParedesAún no hay calificaciones
- Cmponentes y Funcion de MicrogliaDocumento2 páginasCmponentes y Funcion de MicrogliaOK PEROOAún no hay calificaciones
- Neuroplasticidad Neurodesarrllo y Funciones Cognitivo Emocionales 67Documento71 páginasNeuroplasticidad Neurodesarrllo y Funciones Cognitivo Emocionales 67Javier Guevara0% (1)
- Tema 1Documento12 páginasTema 1María Pilar GonzálezAún no hay calificaciones
- 3-Unidad 2 Comp, Neuro 1Documento62 páginas3-Unidad 2 Comp, Neuro 1santiago jofreAún no hay calificaciones
- Sesión N°3Documento30 páginasSesión N°3randol.negocios.07Aún no hay calificaciones
- Neurociencias Actividad #1Documento5 páginasNeurociencias Actividad #1Alejandra Hernandez RodriguezAún no hay calificaciones
- Clase 1 Desarrollo, Maduracioìn y Rganizacioìn Del SNC y SNPDocumento21 páginasClase 1 Desarrollo, Maduracioìn y Rganizacioìn Del SNC y SNPkhendra matusAún no hay calificaciones
- Conducción NerviosaDocumento9 páginasConducción NerviosaYrmaleza de RomeroAún no hay calificaciones
- Ciencias Naturales GRADO 8°Documento7 páginasCiencias Naturales GRADO 8°Yarieth RomeroAún no hay calificaciones
- Tema 1. Niveles Anatomofuncionales de SNCDocumento12 páginasTema 1. Niveles Anatomofuncionales de SNCYeferson CastroAún no hay calificaciones
- Actividad 1 BL 1 Neurofisiología y Neuroanatomía.Documento31 páginasActividad 1 BL 1 Neurofisiología y Neuroanatomía.Ariel GómezAún no hay calificaciones
- NEUROGLÍASDocumento3 páginasNEUROGLÍASFabrizzio Morales ZaldivarAún no hay calificaciones
- Resumen de Todo para El ParcialDocumento11 páginasResumen de Todo para El ParcialMARGY LIZ SUÁREZ CASTELLARAún no hay calificaciones
- Ensayo, NeuronasDocumento7 páginasEnsayo, Neuronasjudith salinasAún no hay calificaciones
- Tejido NerviosoDocumento63 páginasTejido NerviosoyorgeAún no hay calificaciones
- Tejido NerviosoDocumento41 páginasTejido NerviosoItaty JaramilloAún no hay calificaciones
- Guia Sistema Nervioso - Juan A SanchezDocumento12 páginasGuia Sistema Nervioso - Juan A SanchezEvelin FontalvoAún no hay calificaciones
- Neurona, Celula Glia y Potencial de AccionDocumento3 páginasNeurona, Celula Glia y Potencial de AccionJuno CielAún no hay calificaciones
- IntroducciónDocumento13 páginasIntroducciónproyectofrid-toiw08100% (3)
- Actividad IndividualDocumento4 páginasActividad IndividualCamila RuizAún no hay calificaciones
- Clase de NeurofisioDocumento6 páginasClase de Neurofisiosantiago suarezAún no hay calificaciones
- Estc3admulos y Respuestas en El Ser Humano Guc3ada nc2b0 3Documento13 páginasEstc3admulos y Respuestas en El Ser Humano Guc3ada nc2b0 3willy LópezAún no hay calificaciones
- Tejido NerviosoDocumento6 páginasTejido NerviosoAby AguilarAún no hay calificaciones
- Mapa Conceptual 2Documento3 páginasMapa Conceptual 2Dianita Curimilma0% (1)
- PlasticidadDocumento5 páginasPlasticidadChristian HidalgoAún no hay calificaciones
- Evidencia 4 AnatomiaDocumento8 páginasEvidencia 4 AnatomiaJacky CarrilloAún no hay calificaciones
- 3ro de Secundaria Tema 2 SN PDFDocumento6 páginas3ro de Secundaria Tema 2 SN PDFOliver GuarachiAún no hay calificaciones
- Guardado Con Autorrecuperación de Guardado Con Autorrecuperación de INFORMÉ PROYECTIVO DEL TEST DE GOODENOUGHDocumento8 páginasGuardado Con Autorrecuperación de Guardado Con Autorrecuperación de INFORMÉ PROYECTIVO DEL TEST DE GOODENOUGHLida SaldañaAún no hay calificaciones
- Informe de EvaluaciónDocumento1 páginaInforme de EvaluaciónLida SaldañaAún no hay calificaciones
- Actividades Del ESFDocumento1 páginaActividades Del ESFLida SaldañaAún no hay calificaciones
- (AC-S03) Semana 03Documento3 páginas(AC-S03) Semana 03Lida SaldañaAún no hay calificaciones
- Actividades Del ESFDocumento1 páginaActividades Del ESFLida SaldañaAún no hay calificaciones
- Cuestionario de Habilidades de Interacción SocialesDocumento3 páginasCuestionario de Habilidades de Interacción SocialesLida SaldañaAún no hay calificaciones
- Actividades Del ESFDocumento1 páginaActividades Del ESFLida SaldañaAún no hay calificaciones
- 2021 Línea Del Tiempo 2 2021Documento2 páginas2021 Línea Del Tiempo 2 2021Lida SaldañaAún no hay calificaciones
- Analisis de CasosDocumento2 páginasAnalisis de CasosLida SaldañaAún no hay calificaciones
- BiologiaDocumento12 páginasBiologiaLida SaldañaAún no hay calificaciones
- Cómo Revertir Los Juicios LimitantesDocumento1 páginaCómo Revertir Los Juicios LimitantesLida SaldañaAún no hay calificaciones
- El Psicólogo Se Encuentra Revisando Sus Hojas de Sus PacientesDocumento2 páginasEl Psicólogo Se Encuentra Revisando Sus Hojas de Sus PacientesLida SaldañaAún no hay calificaciones
- Registro de Observación No SistematizadaDocumento2 páginasRegistro de Observación No SistematizadaLida Saldaña100% (1)
- (AC-S17) Trabajo Final (TF)Documento13 páginas(AC-S17) Trabajo Final (TF)Lida SaldañaAún no hay calificaciones
- Conducta de SeduccionDocumento1 páginaConducta de SeduccionLida Saldaña100% (1)
- ENTREVISTA SEGUN MARCO TEORICO (Diego)Documento3 páginasENTREVISTA SEGUN MARCO TEORICO (Diego)Lida SaldañaAún no hay calificaciones
- Corto Me TrajeDocumento1 páginaCorto Me TrajeLida SaldañaAún no hay calificaciones
- Corto Me TrajeDocumento1 páginaCorto Me TrajeLida SaldañaAún no hay calificaciones
- Cómo Revertir Los Juicios LimitantesDocumento1 páginaCómo Revertir Los Juicios LimitantesLida SaldañaAún no hay calificaciones
- Corto Me TrajeDocumento1 páginaCorto Me TrajeLida SaldañaAún no hay calificaciones
- 2021 Línea Del Tiempo 2 2021Documento2 páginas2021 Línea Del Tiempo 2 2021Lida SaldañaAún no hay calificaciones
- Libro de Bebes de Lois Bly, AedinDocumento226 páginasLibro de Bebes de Lois Bly, AedinJessica Alejandra Pizarro Morales92% (12)
- 03 Clases de Celulas - Procariotas-Eucariotas PDFDocumento2 páginas03 Clases de Celulas - Procariotas-Eucariotas PDFhortensia100% (1)
- Calendario Gymvirtual Marzo 2024 IntDocumento1 páginaCalendario Gymvirtual Marzo 2024 IntManuela BerrielAún no hay calificaciones
- Atp MioDocumento44 páginasAtp MioPaola TraibelAún no hay calificaciones
- Sesion 6 Mecanica CorporalDocumento31 páginasSesion 6 Mecanica CorporalKarla SilvaAún no hay calificaciones
- La Sombra NegraDocumento7 páginasLa Sombra NegraDavis AvilbarAún no hay calificaciones
- Actividad 1 Comprension LectoraDocumento4 páginasActividad 1 Comprension LectoraPaulina Hernández RiveraAún no hay calificaciones
- Historia Clínica PediátricaDocumento10 páginasHistoria Clínica PediátricaIsabel Chirinos100% (3)
- Libro NAC Tomo 2Documento760 páginasLibro NAC Tomo 2Pamelita Gutierrez Portela85% (13)
- 02-Dolor Toracico en EmergenciaDocumento58 páginas02-Dolor Toracico en EmergenciaLunarcito Jimenezg100% (1)
- Carbohidratos termINADODocumento27 páginasCarbohidratos termINADOFranz CaliAún no hay calificaciones
- Uso de Cafeína en NeonatosDocumento14 páginasUso de Cafeína en NeonatosJannelly HerreraAún no hay calificaciones
- Expo PelvisDocumento44 páginasExpo PelvisNeirod Chaupis MezaAún no hay calificaciones
- Captación de Proteínas en Las MitocondriasDocumento2 páginasCaptación de Proteínas en Las MitocondriasJudy Colmenero50% (2)
- Ciclo Celular Gab 1Documento17 páginasCiclo Celular Gab 1LuzGabrielaSalazarAvilaAún no hay calificaciones
- He Nacido para Verte Sonreir - Santiago LozaDocumento24 páginasHe Nacido para Verte Sonreir - Santiago LozaGabriel Grundig100% (5)
- Cómo Empezar A Estimular - El Sonido de La Hierba Al CrecerDocumento12 páginasCómo Empezar A Estimular - El Sonido de La Hierba Al CrecermatiasAún no hay calificaciones
- Mapa Mental Agujeros Del CraneoDocumento3 páginasMapa Mental Agujeros Del CraneoLourdes Faviana Cueto MonteroAún no hay calificaciones
- Tabla de Sistema TergumetarioDocumento5 páginasTabla de Sistema TergumetarioNandaAún no hay calificaciones
- Sistema Del ComplementoDocumento19 páginasSistema Del ComplementoΛορεηαΣανξεζ0% (1)
- ScyphozoaDocumento21 páginasScyphozoaDaniel AntunezAún no hay calificaciones
- Lesiones Rojo AzuladasDocumento9 páginasLesiones Rojo AzuladasalexanderAún no hay calificaciones
- Hemo Component EsDocumento17 páginasHemo Component EsederAún no hay calificaciones
- Oxitocicos y TocoliticosDocumento3 páginasOxitocicos y TocoliticosHilma VásquezAún no hay calificaciones
- Caso Clinico CiezaDocumento14 páginasCaso Clinico CiezaleslyjanetAún no hay calificaciones
- Informe de LaboratorioDocumento5 páginasInforme de LaboratorioRosseveth Richard Chino HuisaAún no hay calificaciones
- Seminario 1 Dra YUSLENNYDocumento15 páginasSeminario 1 Dra YUSLENNYYuslenny AponteAún no hay calificaciones
- Abordaje Diagnostico de AnemiaDocumento25 páginasAbordaje Diagnostico de AnemiaVictor Fernando Avilez ElizaldeAún no hay calificaciones
- Ciclo de Kresb Usmp IIDocumento34 páginasCiclo de Kresb Usmp IImariajoseAún no hay calificaciones
- Ejercicios-de-Comprension-Lectora-de-Lengua-5º-primaria-Editorial-Anaya (Arrastrado) 2Documento2 páginasEjercicios-de-Comprension-Lectora-de-Lengua-5º-primaria-Editorial-Anaya (Arrastrado) 2ainara.alcalde.gallegoAún no hay calificaciones