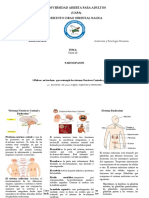
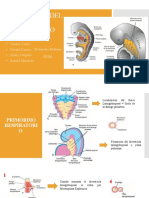
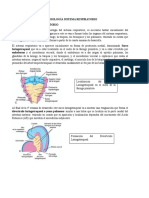
También podría gustarte
- Desarrollo Embrionario Del SNC 15-11-2021Documento5 páginasDesarrollo Embrionario Del SNC 15-11-2021Johomara Arlett Fernandez PrietoAún no hay calificaciones
- Monografia AnatomiaDocumento18 páginasMonografia AnatomiaBeatriz Florian CuevaAún no hay calificaciones
- Capítulo IIDocumento50 páginasCapítulo IINicolle Constanza Reiher MaierAún no hay calificaciones
- Tarea IIIDocumento4 páginasTarea IIIVioleta Caceres De AzaAún no hay calificaciones
- Sistema NerviosoDocumento37 páginasSistema NerviosoDaisy ConchaAún no hay calificaciones
- Elemento1143568 1636568353Documento34 páginasElemento1143568 1636568353Jessica YanonAún no hay calificaciones
- Unidad N 8 Biologia 12 Grado Anatomia y Fisiologia Humana 5Documento24 páginasUnidad N 8 Biologia 12 Grado Anatomia y Fisiologia Humana 5Kris ChavezAún no hay calificaciones
- Resumen NeurofisiologíaDocumento34 páginasResumen NeurofisiologíaVirginia MaitreAún no hay calificaciones
- Ayudantía 3 18 - 08Documento23 páginasAyudantía 3 18 - 08Claudia PizarroAún no hay calificaciones
- Infografía Del Sistema Nervioso CENTRAL Y PERIFERICODocumento2 páginasInfografía Del Sistema Nervioso CENTRAL Y PERIFERICOKATY MARIN QUISPE RODRIGUEZ100% (1)
- PSICOLOGÍA I Guía SEGUNDO PARCIALDocumento17 páginasPSICOLOGÍA I Guía SEGUNDO PARCIALAndriyjrAún no hay calificaciones
- Tarea 2Documento16 páginasTarea 2wellinston quevedoAún no hay calificaciones
- Neuro AnatomiaDocumento85 páginasNeuro AnatomiaRICARDO CAMACHO LANDEROSAún no hay calificaciones
- Infografia Del Sistema Nervioso 152772 Downloable 1036020Documento4 páginasInfografia Del Sistema Nervioso 152772 Downloable 1036020Manita GDAún no hay calificaciones
- Clase 4. Ontogénia y FilogeniaDocumento39 páginasClase 4. Ontogénia y FilogeniaLuciano Paqui GodoyAún no hay calificaciones
- Capitulo 2 Diojani SepulvedaDocumento7 páginasCapitulo 2 Diojani SepulvedaDiojani Sepulveda Terrero100% (1)
- Infografia Del Sistema NerviosoDocumento2 páginasInfografia Del Sistema NerviosoangelicaAún no hay calificaciones
- Anatomia Sistema NerviosoDocumento14 páginasAnatomia Sistema NerviosoJosé Marvin Arriola RodríguezAún no hay calificaciones
- Introducción A La PsicologíaDocumento32 páginasIntroducción A La PsicologíaThyta ShuguliAún no hay calificaciones
- Infografia El Sistema Nervioso y Su EstructuraDocumento1 páginaInfografia El Sistema Nervioso y Su EstructuraNicolle100% (3)
- Sistema NerviosoDocumento7 páginasSistema NerviosoMia DiazAún no hay calificaciones
- Neuropsicologia Dinamica Integral CeciliaDocumento21 páginasNeuropsicologia Dinamica Integral Ceciliachechu.altamirano26Aún no hay calificaciones
- Guia 1 Grado Octavo Ciencias Naturales JCPFDocumento8 páginasGuia 1 Grado Octavo Ciencias Naturales JCPFJUAN CARLOS PONCEAún no hay calificaciones
- Clase 8 Nalto y N BajoDocumento46 páginasClase 8 Nalto y N BajoLeinad IdkAún no hay calificaciones
- Accede A Apuntes, Guías, Libros y Más de Tu Carrera: Infografia-Del-Sistema-Nervioso 1 PagDocumento2 páginasAccede A Apuntes, Guías, Libros y Más de Tu Carrera: Infografia-Del-Sistema-Nervioso 1 PagFlor RsAún no hay calificaciones
- Sistema NeurologicoDocumento16 páginasSistema Neurologicoagueda.ojeda6280Aún no hay calificaciones
- Neurofisiologia UNIDAD 1Documento18 páginasNeurofisiologia UNIDAD 1Alejandra SueldoAún no hay calificaciones
- Cap 7 - TÉJIDO NERVIOSO FINAL (Histologia de Leslie P - Gartner)Documento52 páginasCap 7 - TÉJIDO NERVIOSO FINAL (Histologia de Leslie P - Gartner)Lidia Carolina Rivera Tapia50% (2)
- FIL ADD: Unidad 1Documento45 páginasFIL ADD: Unidad 1Samira Leila SerbaliAún no hay calificaciones
- Generalidades Del Sistema NerviosoDocumento13 páginasGeneralidades Del Sistema NerviosoIngrid VarelaAún no hay calificaciones
- Generalidades de NeuroanatomiaDocumento105 páginasGeneralidades de Neuroanatomiajosue huertaAún no hay calificaciones
- Resumen 1 - Morfologia Del Sistema NerviosoDocumento9 páginasResumen 1 - Morfologia Del Sistema NerviosoSaid Izurieta G.Aún no hay calificaciones
- Apunte de Sistema Nervioso 2022Documento18 páginasApunte de Sistema Nervioso 2022Luli PezziniAún no hay calificaciones
- Tarea 1 de Anatomia y Fisiologia Del Sistema NerviosoDocumento9 páginasTarea 1 de Anatomia y Fisiologia Del Sistema NerviosoRosa Maria De La RosaAún no hay calificaciones
- Unidad 2Documento63 páginasUnidad 2Kiara PavezAún no hay calificaciones
- Cuadro Actividad 2Documento5 páginasCuadro Actividad 2Paula WranetAún no hay calificaciones
- Neuroanatomia 2021Documento7 páginasNeuroanatomia 2021Mauricio MuñozAún no hay calificaciones
- Unidad N°1 Sistema NerviosoDocumento21 páginasUnidad N°1 Sistema NerviosoHolayyo JajjajAún no hay calificaciones
- CURSO DE NEUROANATOMÍA - 1°parcialDocumento44 páginasCURSO DE NEUROANATOMÍA - 1°parcialSTHEFAún no hay calificaciones
- Psicología Parte I Mag. Fresia Vega: Temario Desarrollado deDocumento49 páginasPsicología Parte I Mag. Fresia Vega: Temario Desarrollado deAnalucia Ariadne Tocas MerinoAún no hay calificaciones
- Tarea 2 Anatomina y Fisiologia Del Sistema NerviosoDocumento8 páginasTarea 2 Anatomina y Fisiologia Del Sistema NerviosoEsthefany Acevedo CruzAún no hay calificaciones
- OntogenesisDocumento20 páginasOntogenesisGuadalupe Palacios :3Aún no hay calificaciones
- Sistema LimbicoDocumento59 páginasSistema LimbicoSara CuellarAún no hay calificaciones
- Tarea 1 (Anatomia y Fisiologia Del SN)Documento11 páginasTarea 1 (Anatomia y Fisiologia Del SN)andrixsonAún no hay calificaciones
- Módulo de LecturaDocumento11 páginasMódulo de LecturaALICIA PAMELA RODRIGUEZ ALCANTARAAún no hay calificaciones
- Infografia Sistema NerviosoDocumento1 páginaInfografia Sistema NerviosovanessaAún no hay calificaciones
- Sistema Nervioso Central AnimalDocumento9 páginasSistema Nervioso Central AnimalJesus JimenezAún no hay calificaciones
- Tarea 2 - Nociones Generales de La AnatomiaDocumento5 páginasTarea 2 - Nociones Generales de La Anatomiaruth riveraAún no hay calificaciones
- Generalidades-Del-Sistema-Nervioso UeUDocumento5 páginasGeneralidades-Del-Sistema-Nervioso UeUAmil Diney Ali CadenaAún no hay calificaciones
- Dossier de MarifelDocumento11 páginasDossier de MarifelÎśããč Fx SosaAún no hay calificaciones
- Generalidades Organizador PDFDocumento3 páginasGeneralidades Organizador PDFDIGNÓ H. O.Aún no hay calificaciones
- Generalidades Sistema NerviosoDocumento11 páginasGeneralidades Sistema NerviosoDaniela Villarroel100% (1)
- Genesis NeurobiologíaDocumento5 páginasGenesis NeurobiologíaJean carlos LunaAún no hay calificaciones
- Clase 6 Sistema NerviosoDocumento4 páginasClase 6 Sistema NerviosoFrancesca PascualAún no hay calificaciones
- Plan de Prevención y Control de Patologías de Origen LaboralDocumento19 páginasPlan de Prevención y Control de Patologías de Origen LaboralAlexander Mejia Navarro100% (1)
- Entrega 1 Psicologia Mapa MentalDocumento6 páginasEntrega 1 Psicologia Mapa MentalAuraliz Joly CastroAún no hay calificaciones
- Histologia Sistema Nervioso ContenidoDocumento19 páginasHistologia Sistema Nervioso Contenidovictoria sofiaAún no hay calificaciones
- Guía Grado 9 Sistema NerviosoDocumento5 páginasGuía Grado 9 Sistema Nerviosogustavo herreraAún no hay calificaciones
- Anatomia y Fisiología del Sistema Nervioso IIDe EverandAnatomia y Fisiología del Sistema Nervioso IICalificación: 4 de 5 estrellas4/5 (28)
- Diapositivas Medicina EmbriologíaDocumento19 páginasDiapositivas Medicina EmbriologíaHilaryAún no hay calificaciones
- Arbovirus y Virus Zoonoticos MedicinaDocumento42 páginasArbovirus y Virus Zoonoticos MedicinaHilaryAún no hay calificaciones
- Review Embrio III MedicinaDocumento3 páginasReview Embrio III MedicinaHilaryAún no hay calificaciones
- Apuntes Sistema Nervioso MedicinaDocumento2 páginasApuntes Sistema Nervioso MedicinaHilaryAún no hay calificaciones
- Adaptacioncelular 110813005708 Phpapp01Documento34 páginasAdaptacioncelular 110813005708 Phpapp01Carla L. Castillo MagariñosAún no hay calificaciones
- Sistema Respiratorio Embrio Exposición MedicinaDocumento9 páginasSistema Respiratorio Embrio Exposición MedicinaHilaryAún no hay calificaciones
- Review Sistema Cardiov EmbriologíaDocumento20 páginasReview Sistema Cardiov EmbriologíaHilaryAún no hay calificaciones
- Diversidad Celular en MedicinaDocumento59 páginasDiversidad Celular en MedicinaHilaryAún no hay calificaciones
- Historia Natural Del Lupus E. S. SFDocumento3 páginasHistoria Natural Del Lupus E. S. SFHilary50% (2)
- Declaración de YakartaDocumento11 páginasDeclaración de YakartaCEDOM Virtual Profesor Luis GauthierAún no hay calificaciones
- Sindrome Tourette MedicinaDocumento5 páginasSindrome Tourette MedicinaHilaryAún no hay calificaciones
- Enterobacterias. MedicinaDocumento18 páginasEnterobacterias. MedicinaHilaryAún no hay calificaciones
- Glándulas Anexas Anato P IIDocumento48 páginasGlándulas Anexas Anato P IIHilaryAún no hay calificaciones
- Alma Ata 1978declaracionDocumento3 páginasAlma Ata 1978declaracionLucelia Anahi SantosAún no hay calificaciones
- Grupos ABO SanguíneosDocumento12 páginasGrupos ABO SanguíneosHilaryAún no hay calificaciones
- Generalidades Del Sistema EsqueleticoDocumento90 páginasGeneralidades Del Sistema EsqueleticoJenifer Herrera EscorciaAún no hay calificaciones
- Hemo y Mio BioT II CroteDocumento54 páginasHemo y Mio BioT II CroteHilaryAún no hay calificaciones
- HCG (Gravindex) 2020Documento26 páginasHCG (Gravindex) 2020HilaryAún no hay calificaciones
- Cavidad Oral - Glándulas Salivales Anato P IIDocumento53 páginasCavidad Oral - Glándulas Salivales Anato P IIHilaryAún no hay calificaciones
- Huesos Del NeurocráneoDocumento10 páginasHuesos Del NeurocráneoHilaryAún no hay calificaciones
- Estadistica DescriptivaDocumento88 páginasEstadistica DescriptivaHilaryAún no hay calificaciones
- Generalidades Del Sistema EsqueleticoDocumento20 páginasGeneralidades Del Sistema EsqueleticoHilaryAún no hay calificaciones
- Diapo. Cabeza Ósea 2 Corte Anato. T.Documento154 páginasDiapo. Cabeza Ósea 2 Corte Anato. T.HilaryAún no hay calificaciones
- Práctica de Laboratorio N°9 Síndrome Metabólico.Documento27 páginasPráctica de Laboratorio N°9 Síndrome Metabólico.HilaryAún no hay calificaciones
- Lab UrianalisisDocumento13 páginasLab UrianalisisHilaryAún no hay calificaciones
- Datos y VariablesDocumento18 páginasDatos y VariablesHilaryAún no hay calificaciones
- Lectura 8 - LipoproteínasDocumento23 páginasLectura 8 - LipoproteínasAntonio MartinezAún no hay calificaciones
- Regiones AbdominalDocumento51 páginasRegiones AbdominalHilaryAún no hay calificaciones
- Catedras Bioquimica - LipoproteinasDocumento9 páginasCatedras Bioquimica - Lipoproteinasjoseph48Aún no hay calificaciones
- Crianza de ElefantesDocumento2 páginasCrianza de ElefantesMarco Joe Alegría VásquezAún no hay calificaciones
- Actividad 1.aDocumento2 páginasActividad 1.aDoña BellitaAún no hay calificaciones
- Unidad 11Documento210 páginasUnidad 11JULIAN SANTIAGO CASTIBLANCO GIRALDOAún no hay calificaciones
- Proyecto Ciclo Estral de Las CobayasDocumento15 páginasProyecto Ciclo Estral de Las CobayasAngela Avilez ZambranoAún no hay calificaciones
- Cartel Placenta Previa - Dppni-Mola HidatiformeDocumento3 páginasCartel Placenta Previa - Dppni-Mola HidatiformeStephanie SalazarAún no hay calificaciones
- PLANIFICACION MICROCURRICULAR PROYECTO - 2DO BGU - BT - PROYECTO 2 - Semana 15 A 18Documento11 páginasPLANIFICACION MICROCURRICULAR PROYECTO - 2DO BGU - BT - PROYECTO 2 - Semana 15 A 18alejandrajervezAún no hay calificaciones
- Yondome Wa Iyana Shi Zokusei Majutsushi - Arco 09 (Incompleto)Documento524 páginasYondome Wa Iyana Shi Zokusei Majutsushi - Arco 09 (Incompleto)gwhrsfdmkrAún no hay calificaciones
- Guia U3 BiologíaDocumento12 páginasGuia U3 Biologíahariz inversionesAún no hay calificaciones
- 2da Evaluacion 2020 SarayDocumento4 páginas2da Evaluacion 2020 Sarayyemilys aldanaAún no hay calificaciones
- Ciclo CelularDocumento7 páginasCiclo CelularMichell ReyesAún no hay calificaciones
- Final de PropaDocumento2 páginasFinal de PropaAlexis Pool Zuñiga HuertasAún no hay calificaciones
- Semana 7 Aparato Reproductor FemeninoDocumento35 páginasSemana 7 Aparato Reproductor FemeninoVíctor game YTAún no hay calificaciones
- Historia PsicosocialDocumento2 páginasHistoria PsicosocialClau ́s ChannelAún no hay calificaciones
- Práctica BiodiversidadDocumento24 páginasPráctica BiodiversidadCarolina UrazánAún no hay calificaciones
- Multiplicacion de Las PlantasDocumento28 páginasMultiplicacion de Las PlantasSteice Gabriela Lescano OlivaAún no hay calificaciones
- 1er Parcial Biologia 2Documento5 páginas1er Parcial Biologia 2Alan Velazquez Shade100% (2)
- Callo (Biología)Documento4 páginasCallo (Biología)Stephanie MirandaAún no hay calificaciones
- Informe Seleccion de ReproductoresDocumento15 páginasInforme Seleccion de ReproductoresJosue VeraAún no hay calificaciones
- Guia No 7 Observacion y Reconocimiento de Hongos 2016-1Documento15 páginasGuia No 7 Observacion y Reconocimiento de Hongos 2016-1rossmiAún no hay calificaciones
- La Perdida LemuriaDocumento19 páginasLa Perdida Lemuriaviolett1111100% (2)
- Programacion Tesela Biologia y Geologia 1 BACH Comunitat ValencianaDocumento55 páginasProgramacion Tesela Biologia y Geologia 1 BACH Comunitat ValencianajfzepolAún no hay calificaciones
- Ensayo Metosis Meiosis GametogenesisDocumento4 páginasEnsayo Metosis Meiosis GametogenesisroselysAún no hay calificaciones
- GametogenesisDocumento11 páginasGametogenesisVanessa Rodriguez OlmosAún no hay calificaciones
- OTECDocumento219 páginasOTECClaudia ValenzuelaAún no hay calificaciones
- Meiosis IDocumento16 páginasMeiosis IOrtodoncia InerAún no hay calificaciones
- Aparato Reproductor Humano para Sexto de PrimariaDocumento8 páginasAparato Reproductor Humano para Sexto de PrimariaJoel Paz AguilarAún no hay calificaciones
- Aparato GenitalDocumento51 páginasAparato GenitalJOSELIN MILAGROS RAMOS HUANCAAún no hay calificaciones
- Embriologia Experimental PDFDocumento9 páginasEmbriologia Experimental PDFbixby51Aún no hay calificaciones
- Presentacion Introduccion A La Produccion Animal1Documento14 páginasPresentacion Introduccion A La Produccion Animal1Mariangel SalazarAún no hay calificaciones
- Tarea 1 - Ríos Vásquez Paola MiladyDocumento3 páginasTarea 1 - Ríos Vásquez Paola MiladyJophnny GutierrezAún no hay calificaciones