También podría gustarte
- Las cadenas fisiológicas (Tomo II): La cintura pélvica y el miembro inferiorDe EverandLas cadenas fisiológicas (Tomo II): La cintura pélvica y el miembro inferiorCalificación: 4.5 de 5 estrellas4.5/5 (10)
- Fisiología del ejercicio físico y del entrenamientoDe EverandFisiología del ejercicio físico y del entrenamientoCalificación: 4.5 de 5 estrellas4.5/5 (8)
- Las cadenas fisiológicas (Tomo VII): La cadena visceral. Tórax, garganta y bocaDe EverandLas cadenas fisiológicas (Tomo VII): La cadena visceral. Tórax, garganta y bocaAún no hay calificaciones
- Las cadenas fisiológicas (Tomo VI): La cadena visceral, Abdomen - PelvisDe EverandLas cadenas fisiológicas (Tomo VI): La cadena visceral, Abdomen - PelvisCalificación: 4 de 5 estrellas4/5 (2)
- Solucionario Fisica Universitaria de SerasDocumento18 páginasSolucionario Fisica Universitaria de SerasYoselin Samay Cuero SinisterraAún no hay calificaciones
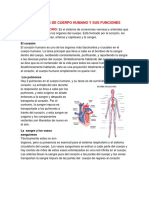
- 11 Sistemas de Cuerpo Humano y Sus Funciones y OrganosDocumento11 páginas11 Sistemas de Cuerpo Humano y Sus Funciones y OrganosMaría De Los Angeles100% (8)
- Adap Musc EjercicioDocumento6 páginasAdap Musc EjercicioJareth AstorgaAún no hay calificaciones
- Adaptacion Muscular Al EjercicioDocumento6 páginasAdaptacion Muscular Al EjercicioAnapatriciaAún no hay calificaciones
- Fibras Mucsulares PDFDocumento10 páginasFibras Mucsulares PDFLindaAún no hay calificaciones
- Observaciones Sobre Los Distintos Tipos de Células o Fibras Musculares A Nivel Metabólico, Molecular y Su Rendimiento en La Actividad FísicaDocumento10 páginasObservaciones Sobre Los Distintos Tipos de Células o Fibras Musculares A Nivel Metabólico, Molecular y Su Rendimiento en La Actividad FísicaLudmila CaseresAún no hay calificaciones
- Tema 7 Fisiología de La Fibra MuscularDocumento27 páginasTema 7 Fisiología de La Fibra MuscularJordiAún no hay calificaciones
- Ra XimhaiDocumento17 páginasRa XimhaiKenneth FernándezAún no hay calificaciones
- Ensayo Invetigativo Fisiologia Del EsfuerzoDocumento7 páginasEnsayo Invetigativo Fisiologia Del EsfuerzoRuth VelascoAún no hay calificaciones
- TALLER No4 FC MEDICINADocumento3 páginasTALLER No4 FC MEDICINAAlejandro Ortega MuñozAún no hay calificaciones
- Respuesta Muscular Al EjercicioDocumento8 páginasRespuesta Muscular Al EjercicioaleajandroAún no hay calificaciones
- Fisiología MuscularDocumento4 páginasFisiología MuscularMari CarmenAún no hay calificaciones
- Tipos de Fibras Musculares Características y FunciónDocumento8 páginasTipos de Fibras Musculares Características y Funciónyelitza marinAún no hay calificaciones
- BoffiDocumento5 páginasBoffiEL HIJO DE DIOSAún no hay calificaciones
- Tipos de Fibras Musculares PDFDocumento4 páginasTipos de Fibras Musculares PDFCadHomeAún no hay calificaciones
- 2.tipos de Fibra Muscular EsqueleticoDocumento4 páginas2.tipos de Fibra Muscular EsqueleticoNavia BreeizyAún no hay calificaciones
- Dialnet ElectroforesisEnGelDePoliacrilamidaSDSComoHerramie 6020409 PDFDocumento20 páginasDialnet ElectroforesisEnGelDePoliacrilamidaSDSComoHerramie 6020409 PDFYury Daniela Giraldo PajoyAún no hay calificaciones
- Silabo de Fisiologia FMH 1Documento19 páginasSilabo de Fisiologia FMH 1silverAún no hay calificaciones
- PM 4099Documento21 páginasPM 4099Elkin Dario Antolinez MuñozAún no hay calificaciones
- PHBP 02Documento395 páginasPHBP 02viviana.nurse.2021Aún no hay calificaciones
- Dialnet FisiologiaEnElEntrenamientoDeLaAptitudFisicaMuscul 5289103Documento8 páginasDialnet FisiologiaEnElEntrenamientoDeLaAptitudFisicaMuscul 5289103MARÍA CRISTINA CHICA MARULANDAAún no hay calificaciones
- Propiedades Fisiologicas Del Musculo EsqueleticoDocumento3 páginasPropiedades Fisiologicas Del Musculo EsqueleticoMayumi Torres CastilloAún no hay calificaciones
- Tema 2013 El Sistema MuscularDocumento5 páginasTema 2013 El Sistema MuscularangelgadAún no hay calificaciones
- Unidad III Bases Fisiologicas Del Movimiento HumanoDocumento26 páginasUnidad III Bases Fisiologicas Del Movimiento HumanoAdyaf Sagev83% (6)
- Bioquimica II-practica 09 FaltaDocumento31 páginasBioquimica II-practica 09 FaltaChabelly Darling Calderón MarínAún no hay calificaciones
- TEMA 03 - Metabolismo Musculo Esquelético y Actividad MotoraDocumento17 páginasTEMA 03 - Metabolismo Musculo Esquelético y Actividad MotoraDavid García MartínezAún no hay calificaciones
- Sistema MuscularDocumento9 páginasSistema MuscularJorge RodríguezAún no hay calificaciones
- ZoteroDocumento5 páginasZoteroKarlaAún no hay calificaciones
- Apuntes de Fisiología II Víctor Illera PDFDocumento46 páginasApuntes de Fisiología II Víctor Illera PDFJuan CardenasAún no hay calificaciones
- Fisilogia Humana J Taller Corte 1 Brayan SarayDocumento5 páginasFisilogia Humana J Taller Corte 1 Brayan SarayBRAYAN STIWENS SARAY GORDILLOAún no hay calificaciones
- mTORC1 in The Control of Myogenesis and Adult Skeletal Muscle MassDocumento20 páginasmTORC1 in The Control of Myogenesis and Adult Skeletal Muscle MassJulio E Benitez MontesAún no hay calificaciones
- Jorge Cancino Fisiología Del EjercicioDocumento34 páginasJorge Cancino Fisiología Del EjercicioKaren Torres PantojaAún no hay calificaciones
- Propiedades Fisiológicas Del Músculo EsqueléticoDocumento3 páginasPropiedades Fisiológicas Del Músculo EsqueléticoAli Yamilé Camizan Cunias50% (2)
- 9 - PRÁCTICO N Tejido MuscularDocumento1 página9 - PRÁCTICO N Tejido MuscularKalendra MiniaturasAún no hay calificaciones
- Mejor RespuestaDocumento15 páginasMejor RespuestaRigoberto Jimenez GonzalezAún no hay calificaciones
- Programa de Fisiologia Cartas Descriptivas de Todo Fisiologia para Medicina FO 101 1Documento85 páginasPrograma de Fisiologia Cartas Descriptivas de Todo Fisiologia para Medicina FO 101 1GabrielaMejíaValladaresAún no hay calificaciones
- Fisica BiologicaDocumento17 páginasFisica BiologicaGitana MorenaAún no hay calificaciones
- FISIOLOGÍADocumento16 páginasFISIOLOGÍARosarioAún no hay calificaciones
- Biomecanica de MusculoDocumento20 páginasBiomecanica de MusculoHumphrey Cruzado Catalan SandovalAún no hay calificaciones
- Silabus de Fisiologia - Medicina Humana - 2010-I - (1) - Universidad ...Documento19 páginasSilabus de Fisiologia - Medicina Humana - 2010-I - (1) - Universidad ...Jean Paul BalarezoAún no hay calificaciones
- Actividad Individual CarnicosDocumento3 páginasActividad Individual CarnicosLuisa Sanchez VillalobosAún no hay calificaciones
- Sesi 02Documento4 páginasSesi 02manson msmsAún no hay calificaciones
- Practica Anatomia Del CorazonDocumento16 páginasPractica Anatomia Del CorazonDaniel AlbertoAún no hay calificaciones
- Bioquímica de La CarneDocumento22 páginasBioquímica de La CarneKaren YamaliAún no hay calificaciones
- Bioenergética de Las Fibras Musculares y Ejercicio by José López Chicharro Almudena Fernández VaqueroDocumento61 páginasBioenergética de Las Fibras Musculares y Ejercicio by José López Chicharro Almudena Fernández VaqueroGustavo MartinezAún no hay calificaciones
- 1er TemaDocumento10 páginas1er TemaEscobedo FreddyAún no hay calificaciones
- Respuesta Fisiológica Del Tejido Conjuntivo de Músculos y Tendones Tras La Aplicación de Los Agentes FísicosDocumento7 páginasRespuesta Fisiológica Del Tejido Conjuntivo de Músculos y Tendones Tras La Aplicación de Los Agentes FísicosThiareGeraldineAún no hay calificaciones
- Tejido. MuscularDocumento11 páginasTejido. MuscularFreidy GarciaAún no hay calificaciones
- Articulo MúsculoDocumento13 páginasArticulo MúsculoJuan Camilo MenesesAún no hay calificaciones
- Plasticidad Muscular y Fibras Hi ÜbridasDocumento11 páginasPlasticidad Muscular y Fibras Hi ÜbridasCarolina Lissette Poblete BeltránAún no hay calificaciones
- GD1-Entrenamiento PersonalizadoDocumento33 páginasGD1-Entrenamiento PersonalizadoCriszehro ReviewerAún no hay calificaciones
- ANATOMÍADocumento2 páginasANATOMÍAMateo EnriqueAún no hay calificaciones
- Ensayo de Tejido MuscularDocumento10 páginasEnsayo de Tejido MuscularCelia QuintalAún no hay calificaciones
- Cronograma 2023 Hasta 1ra PDocumento7 páginasCronograma 2023 Hasta 1ra PGabi JaraAún no hay calificaciones
- Bioquímica Del EntrenamientoDocumento4 páginasBioquímica Del EntrenamientopedroAún no hay calificaciones
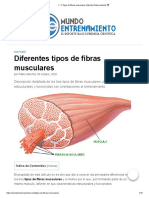
- 3 Tipos de Fibras Musculares - Mundo Entrenamiento ?Documento8 páginas3 Tipos de Fibras Musculares - Mundo Entrenamiento ?abner romavi vargas uhAún no hay calificaciones
- Material de Estudio. NeurofisiologíaDocumento29 páginasMaterial de Estudio. NeurofisiologíaMarcela LeivaAún no hay calificaciones
- Los 50 Inventos TecnologicosDocumento12 páginasLos 50 Inventos TecnologicosYoselin Samay Cuero SinisterraAún no hay calificaciones
- LógicaMatemática PDFDocumento81 páginasLógicaMatemática PDFYoselin Samay Cuero SinisterraAún no hay calificaciones
- Perfil Logistico de Ecuador PDFDocumento10 páginasPerfil Logistico de Ecuador PDFYoselin Samay Cuero SinisterraAún no hay calificaciones
- Proceso Exportador de Empresas Santandereanas de Calzado A Ecuador Y VenezuelaDocumento18 páginasProceso Exportador de Empresas Santandereanas de Calzado A Ecuador Y VenezuelaYoselin Samay Cuero SinisterraAún no hay calificaciones
- Caso Clinico Adenopatia y FimosisDocumento23 páginasCaso Clinico Adenopatia y FimosisYoselin Samay Cuero SinisterraAún no hay calificaciones
- Proyecto Final de Investigación Técnicas de InvestigaciónDocumento40 páginasProyecto Final de Investigación Técnicas de InvestigaciónFatima TorresAún no hay calificaciones
- Resumen Area VozDocumento27 páginasResumen Area Vozmarcela riqueroAún no hay calificaciones
- Modulo II Clase II CreatividadDocumento20 páginasModulo II Clase II CreatividadFERNANDA MARGARITA A RIVERA LOPEZAún no hay calificaciones
- Presentación Facilitador en Diplomado 8 de Diciembre de 2018 CONALEP HoyDocumento15 páginasPresentación Facilitador en Diplomado 8 de Diciembre de 2018 CONALEP HoyMum Gay Lázaro CárdenasAún no hay calificaciones
- Sueño, Memoria y AprendizajeDocumento3 páginasSueño, Memoria y AprendizajeClaudia Araujo CaviedesAún no hay calificaciones
- Normativa AshraeDocumento2 páginasNormativa AshraeCruz Erwin100% (1)
- 16PF SistematizaciónDocumento4 páginas16PF SistematizaciónCamilaAún no hay calificaciones
- CuttingDocumento1 páginaCuttingAltamirano JoséAún no hay calificaciones
- Presentacion Cambio Climatico en Colombia.Documento27 páginasPresentacion Cambio Climatico en Colombia.Jessi CaAún no hay calificaciones
- Diez Esferas de Acción para Poner Fin A La Violencia en La EscuelaDocumento7 páginasDiez Esferas de Acción para Poner Fin A La Violencia en La EscuelaJose GuerraAún no hay calificaciones
- TESISDocumento95 páginasTESISmartha villegasAún no hay calificaciones
- Educación Física - Semana 27Documento3 páginasEducación Física - Semana 27Adriana Díaz FariasAún no hay calificaciones
- Nutrición y Eclerodermia. Dña Carmen Urzola PDFDocumento20 páginasNutrición y Eclerodermia. Dña Carmen Urzola PDFD Estefanía ZabalaAún no hay calificaciones
- Laceado BrasileroDocumento4 páginasLaceado BrasileroAlejandra Loli HernándezAún no hay calificaciones
- Epidemiología BoliviaDocumento2 páginasEpidemiología BoliviaFranklin Denis Mamani CarpioAún no hay calificaciones
- Banco de SangreDocumento16 páginasBanco de SangreSOPORTE TECNICO MEBCOAún no hay calificaciones
- UVD-TSST - RobayoAguirreEduarLeonardo - 2018Documento54 páginasUVD-TSST - RobayoAguirreEduarLeonardo - 2018Edwin ArteagaAún no hay calificaciones
- Informe Sobre Desarrollo Humano 2001. Naciones UnidasDocumento279 páginasInforme Sobre Desarrollo Humano 2001. Naciones UnidasPedro OlivaAún no hay calificaciones
- SANIDADDocumento13 páginasSANIDADjose colmenarezAún no hay calificaciones
- Peeling FacialDocumento8 páginasPeeling FacialAndrea TorresAún no hay calificaciones
- Historia Clinica GLADYS YOLANDA BALANTA CABEZASDocumento2 páginasHistoria Clinica GLADYS YOLANDA BALANTA CABEZASgladysita1974Aún no hay calificaciones
- Cartilla QuimicosDocumento28 páginasCartilla QuimicosgermanAún no hay calificaciones
- Ruta Formativa Actividad Colaborativa Psicologia Clínica y de La SaludDocumento19 páginasRuta Formativa Actividad Colaborativa Psicologia Clínica y de La SaludSwimwear HeavenAún no hay calificaciones
- Significado Maternidad AdolescenteDocumento203 páginasSignificado Maternidad Adolescentejohnx820Aún no hay calificaciones
- Administración de La Enfermería - Modelos de EnfermeríaDocumento6 páginasAdministración de La Enfermería - Modelos de EnfermeríaK. VelizAún no hay calificaciones
- FORMATO DE INFORME SOCIAL en Word para DiligenciarDocumento8 páginasFORMATO DE INFORME SOCIAL en Word para DiligenciarErnesto BriceñoAún no hay calificaciones
- Experiencia de Aprendizaje NDocumento14 páginasExperiencia de Aprendizaje NNAILEA ZUMAETA MACUYAMAAún no hay calificaciones
- Invecciones Urinarias en El: EmbarazoDocumento6 páginasInvecciones Urinarias en El: EmbarazoJhonyHinojosaAún no hay calificaciones
- 1.1 Concepto de Electroterapia. Efectos Fisicoquímicos y FisiológicosDocumento66 páginas1.1 Concepto de Electroterapia. Efectos Fisicoquímicos y FisiológicosMalonda 17Aún no hay calificaciones