También podría gustarte
- Mesozoico Presentación GabrielDocumento54 páginasMesozoico Presentación GabrielgabrielAún no hay calificaciones
- La Era Mesozoica PDFDocumento5 páginasLa Era Mesozoica PDFJoaquin Poblet JimenezAún no hay calificaciones
- ELECTROLISISDocumento8 páginasELECTROLISISANDREA NICOLE TOLEDO CAZASAún no hay calificaciones
- Clasificación Macroscopica Rocas IgneasDocumento1 páginaClasificación Macroscopica Rocas IgneasSebastián BlackburnAún no hay calificaciones
- Extinción masiva: ¿Realmente hay un 100% de posibilidades de que todas las especies se extingan?De EverandExtinción masiva: ¿Realmente hay un 100% de posibilidades de que todas las especies se extingan?Aún no hay calificaciones
- Geologia de La Mina AntaminaDocumento23 páginasGeologia de La Mina AntaminaShirley Fioreli Espíritu OrtizAún no hay calificaciones
- 107-Tesis-Validacion y Actualizacion Del Modelo Geologico de Los Yacimientos T MFA-33 y T MFA-2Documento110 páginas107-Tesis-Validacion y Actualizacion Del Modelo Geologico de Los Yacimientos T MFA-33 y T MFA-2Pepe Med TolAún no hay calificaciones
- M6C3 PPT ExtincionesDocumento85 páginasM6C3 PPT ExtincionesRaúl QuirogaAún no hay calificaciones
- Extinciones InfoDocumento13 páginasExtinciones InfoRosmary Vanessa Robles RinconAún no hay calificaciones
- Lectura. Extinción y Radiación 2023Documento4 páginasLectura. Extinción y Radiación 2023Alex WheelerAún no hay calificaciones
- Lectura. Extinción y Radiación 2021Documento5 páginasLectura. Extinción y Radiación 2021Antonio Gonzalez VazquezAún no hay calificaciones
- Formato Sesión de Aprendizaje 2023 (Sesion 72)Documento2 páginasFormato Sesión de Aprendizaje 2023 (Sesion 72)tilin 22Aún no hay calificaciones
- El Hombre y Las ExtincionesDocumento3 páginasEl Hombre y Las ExtincionesAnonymous XC8TkOuvLVAún no hay calificaciones
- Estromatolito PDFDocumento14 páginasEstromatolito PDFfernandoAún no hay calificaciones
- CambricoDocumento11 páginasCambricojean claudeAún no hay calificaciones
- Linea Del Tiempo ExtincionesDocumento5 páginasLinea Del Tiempo ExtincionesJosé Jesús Díaz ToscanoAún no hay calificaciones
- Presentacion Era Mesozoica - FinalDocumento46 páginasPresentacion Era Mesozoica - FinalAlvin Vieras100% (1)
- BIOESTRATIGRAFÍADocumento4 páginasBIOESTRATIGRAFÍASophia Cristina Nuñez VerdugoAún no hay calificaciones
- Extinsiones MasivasDocumento3 páginasExtinsiones MasivasKatherine Natalia Buitrago VelandiaAún no hay calificaciones
- Las Eras y PeriodosDocumento4 páginasLas Eras y Periodoserlin_corderoAún no hay calificaciones
- Qué Es La ExtinciónDocumento6 páginasQué Es La ExtinciónantonioAún no hay calificaciones
- Ecologia III Extinción - 124315Documento31 páginasEcologia III Extinción - 124315cado62640Aún no hay calificaciones
- ExtincionesDocumento18 páginasExtincionesAstro AnulAún no hay calificaciones
- IctiosaurioDocumento11 páginasIctiosaurioDaniel Felipe Morales RuedaAún no hay calificaciones
- La Gran Extinción Del Límite K PG en El FlyschDocumento4 páginasLa Gran Extinción Del Límite K PG en El FlyschEmilio AlcainAún no hay calificaciones
- La Era MesozoicaDocumento3 páginasLa Era MesozoicaAriel MartínezAún no hay calificaciones
- El MesozoicoDocumento7 páginasEl Mesozoicoalvaroaydin100% (1)
- ExtincionesDocumento18 páginasExtincionesRodrigo Felix Flores MelendresAún no hay calificaciones
- 01 - Primera Gran ExtinciónDocumento17 páginas01 - Primera Gran ExtinciónJesúsAún no hay calificaciones
- Las Eras GeológicasDocumento14 páginasLas Eras GeológicasTavo VIlla CAún no hay calificaciones
- Taller Origen de La Fauna SudamericanaDocumento3 páginasTaller Origen de La Fauna SudamericanaClara Daniela Charry Marin100% (1)
- Extincion Del PermicoDocumento8 páginasExtincion Del PermicoMax Chavez ChicomaAún no hay calificaciones
- Fauna y Flora en El Periodo CretácicoDocumento3 páginasFauna y Flora en El Periodo CretácicoJOANA RICO MILLANAún no hay calificaciones
- Clase Teórica VI Evolución (2º Parte)Documento38 páginasClase Teórica VI Evolución (2º Parte)Evelyn RauAún no hay calificaciones
- García Ochoa Carmen M14S3 ErasgeologicasDocumento2 páginasGarcía Ochoa Carmen M14S3 ErasgeologicasLilian Chel GuerreroAún no hay calificaciones
- Tríptico de ExtincionesDocumento2 páginasTríptico de ExtincionesMaximo MonoAún no hay calificaciones
- La Era MesozoicaDocumento5 páginasLa Era MesozoicaHelena RivaAún no hay calificaciones
- Álbum de Clasificación de Compuestos QuímicosDocumento10 páginasÁlbum de Clasificación de Compuestos QuímicosShouAún no hay calificaciones
- La Ciencia de La Evolución-17-22-1Documento6 páginasLa Ciencia de La Evolución-17-22-1Johanna GrandaAún no hay calificaciones
- Práctica 6-Microfósiles CalcareosDocumento7 páginasPráctica 6-Microfósiles CalcareosMicroalgae PeruAún no hay calificaciones
- Tema Xi Geología HistóricaDocumento5 páginasTema Xi Geología Históricajaime aguilera ruedaAún no hay calificaciones
- ErasDocumento17 páginasErasCristal BonillaAún no hay calificaciones
- Guía IV Ciencias Sociales SextoDocumento8 páginasGuía IV Ciencias Sociales SextoGanimedes999Aún no hay calificaciones
- El Registro Paleobotanico Y El Transito Permico TriasicoDocumento11 páginasEl Registro Paleobotanico Y El Transito Permico TriasicoRobinsón InfanteAún no hay calificaciones
- Ensayo Evolución TrilobitesDocumento1 páginaEnsayo Evolución Trilobitesdanillorente19Aún no hay calificaciones
- La Mayor Extinción Biológica ConocidaDocumento8 páginasLa Mayor Extinción Biológica ConocidajuanAún no hay calificaciones
- Organic Scrapbook Illustrated Ancient Greece History PresentationDocumento30 páginasOrganic Scrapbook Illustrated Ancient Greece History PresentationJulian PatlaniAún no hay calificaciones
- Evolucion de La VidaDocumento3 páginasEvolucion de La VidaAnthony GomezAún no hay calificaciones
- La Sexta Extincion MasivaDocumento7 páginasLa Sexta Extincion MasivajorgeAún no hay calificaciones
- El Cuartenario Biologia 3 3Documento8 páginasEl Cuartenario Biologia 3 3Benjamin MirandaAún no hay calificaciones
- Carbonifero InferiorDocumento5 páginasCarbonifero InferiorFlosfyAún no hay calificaciones
- Eras Geológicas.Documento4 páginasEras Geológicas.Montserrat GonzalezAún no hay calificaciones
- Eras GeológicasDocumento25 páginasEras GeológicasVaclasofnarcisaAún no hay calificaciones
- Victoria 1Documento16 páginasVictoria 1Araceli NavAún no hay calificaciones
- Actividad en Clase 9Documento3 páginasActividad en Clase 9Lilian BarbozaAún no hay calificaciones
- Las 5 Extinciones en MasaDocumento6 páginasLas 5 Extinciones en MasaAnonymous KD467FCD5Aún no hay calificaciones
- Descripción de Los Factores de Tipo Geológico Que Dieron Origen A La Formación de La Tierra y Su EvoluciónDocumento8 páginasDescripción de Los Factores de Tipo Geológico Que Dieron Origen A La Formación de La Tierra y Su EvoluciónAndres AhmadineyadAún no hay calificaciones
- 0da27a Las Eras Geologicas 2Documento10 páginas0da27a Las Eras Geologicas 2Dawud CespedezAún no hay calificaciones
- 0da27a Las Eras Geologicas 1Documento10 páginas0da27a Las Eras Geologicas 1Dawud CespedezAún no hay calificaciones
- Eras GeologicasDocumento11 páginasEras GeologicasKARLA ISABEL ERAS ATARIHUANAAún no hay calificaciones
- Expocicion CambricoDocumento8 páginasExpocicion CambricoCamilo HernandezAún no hay calificaciones
- Los progresos del hombre y los riesgos de su autodestrucciónDe EverandLos progresos del hombre y los riesgos de su autodestrucciónAún no hay calificaciones
- Libélulas y caballitos del diablo del departamento del Meta, ColombiaDe EverandLibélulas y caballitos del diablo del departamento del Meta, ColombiaAún no hay calificaciones
- Rapidez Del Cambio de La Temperatura Del PlanetaDocumento14 páginasRapidez Del Cambio de La Temperatura Del PlanetaANDREA NICOLE TOLEDO CAZASAún no hay calificaciones
- BoliviaDocumento4 páginasBoliviaANDREA NICOLE TOLEDO CAZASAún no hay calificaciones
- Sismica 3DDocumento3 páginasSismica 3DANDREA NICOLE TOLEDO CAZASAún no hay calificaciones
- Convocatoria Al CampeonatoDocumento4 páginasConvocatoria Al CampeonatoANDREA NICOLE TOLEDO CAZASAún no hay calificaciones
- Sistema de Rotación COMPRDocumento36 páginasSistema de Rotación COMPRANDREA NICOLE TOLEDO CAZASAún no hay calificaciones
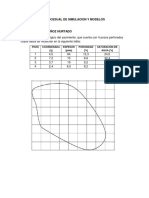
- Carlos Junior Muñoz HuratadoDocumento8 páginasCarlos Junior Muñoz HuratadoANDREA NICOLE TOLEDO CAZASAún no hay calificaciones
- Borja SMDocumento179 páginasBorja SMANDREA NICOLE TOLEDO CAZASAún no hay calificaciones
- TEMA 3 Balance de Materiales en Reservorios de Petróleo (Avance Final 5-Oct-2021)Documento29 páginasTEMA 3 Balance de Materiales en Reservorios de Petróleo (Avance Final 5-Oct-2021)ANDREA NICOLE TOLEDO CAZASAún no hay calificaciones
- Clase 8 Modelos de Inyeccion de Agua 2Documento8 páginasClase 8 Modelos de Inyeccion de Agua 2ANDREA NICOLE TOLEDO CAZASAún no hay calificaciones
- Primer Parcial Simulación y Modelos (Andrea Nicole Toledo Cazas)Documento6 páginasPrimer Parcial Simulación y Modelos (Andrea Nicole Toledo Cazas)ANDREA NICOLE TOLEDO CAZASAún no hay calificaciones
- Freddy Calvi Tarea 1Documento5 páginasFreddy Calvi Tarea 1ANDREA NICOLE TOLEDO CAZASAún no hay calificaciones
- Configuración de PerfilDocumento3 páginasConfiguración de PerfilANDREA NICOLE TOLEDO CAZASAún no hay calificaciones
- Album AnatomiaDocumento20 páginasAlbum AnatomiaANDREA NICOLE TOLEDO CAZASAún no hay calificaciones
- Anatomia 1Documento28 páginasAnatomia 1ANDREA NICOLE TOLEDO CAZASAún no hay calificaciones
- Fallas Geologicas NormalesDocumento3 páginasFallas Geologicas NormalesAndres CuaránAún no hay calificaciones
- Info UnwdgeDocumento15 páginasInfo UnwdgeHebnerAlbertCarhuapomaRivasAún no hay calificaciones
- Diferencia Entre Ondas P y S - Ondas Sísmicas - Diferencias - InfoDocumento1 páginaDiferencia Entre Ondas P y S - Ondas Sísmicas - Diferencias - Infoalvaro.bajoAún no hay calificaciones
- Límites Transformantes de PlacasDocumento10 páginasLímites Transformantes de PlacasJ. Yahir RojasAún no hay calificaciones
- Mapa Geologico de SN-WDocumento1 páginaMapa Geologico de SN-WWalther ChaupiAún no hay calificaciones
- ODocumento5 páginasOKeyla Dzul MayAún no hay calificaciones
- Paragenesis 2020Documento2 páginasParagenesis 2020Gabriela RojasAún no hay calificaciones
- Estabilidad de Taludes en Suelos y RocasDocumento72 páginasEstabilidad de Taludes en Suelos y RocasJUNIOR CONSTANTINO ESPINOZA HUAYRAAún no hay calificaciones
- Porfidos Cupríferos de ChileDocumento3 páginasPorfidos Cupríferos de ChilegonzaloAún no hay calificaciones
- Caballero - T1 - Trabajo SustentadoDocumento21 páginasCaballero - T1 - Trabajo SustentadoAldo Edson Caballero Rodriguez100% (1)
- TEMA N°2 1er Año GFMDocumento10 páginasTEMA N°2 1er Año GFMsilvia cocconcelliAún no hay calificaciones
- Informe Salida A QuirioDocumento27 páginasInforme Salida A QuirioJAMBER SCOTT LUQUE RIVERAAún no hay calificaciones
- 4º Curso A Distancia "Hidrogeología Fundamental"Documento6 páginas4º Curso A Distancia "Hidrogeología Fundamental"Fundación FCIHSAún no hay calificaciones
- Modelo de Los Macizos RocososDocumento33 páginasModelo de Los Macizos RocososAlex Yvan SalcedoAún no hay calificaciones
- 20000131-16-22-48 Ley 2049Documento8 páginas20000131-16-22-48 Ley 2049groverVILLEGASAún no hay calificaciones
- Conociendo El Volcán Arenal - Afiche RSN PDFDocumento1 páginaConociendo El Volcán Arenal - Afiche RSN PDFgjsotoAún no hay calificaciones
- Presentacion de Origen Del PetroleoDocumento68 páginasPresentacion de Origen Del Petroleoana100% (1)
- Tesis Optimización de Producción PetróleosDocumento188 páginasTesis Optimización de Producción Petróleosbrayant duran100% (1)
- Taller Estabilidad SaharaDocumento6 páginasTaller Estabilidad SaharaSaradaniela RicoAún no hay calificaciones
- Silabo PetrologiaDocumento7 páginasSilabo PetrologiaNarcisoAún no hay calificaciones
- Fenomeno MonterreyDocumento13 páginasFenomeno MonterreyTrino QuevedoAún no hay calificaciones
- Informe Geologia 1Documento29 páginasInforme Geologia 1Marianyela CholesAún no hay calificaciones
- Terremotos Ecuador ResumenDocumento65 páginasTerremotos Ecuador ResumenDaniel Tapia MarcosAún no hay calificaciones
- ArenqueDocumento18 páginasArenqueyeroneyAún no hay calificaciones
- Plan de Manejo Ambiental Pavimentación La Coquerita y Alicante PDFDocumento50 páginasPlan de Manejo Ambiental Pavimentación La Coquerita y Alicante PDFgony12Aún no hay calificaciones
- Introducción Final Ontong Java PlateauDocumento2 páginasIntroducción Final Ontong Java PlateauJose GiraldoAún no hay calificaciones
- Mineral GalenaDocumento22 páginasMineral Galenalizeth vargasAún no hay calificaciones