También podría gustarte
- Farmacos IndicadosDocumento8 páginasFarmacos IndicadosDanielAún no hay calificaciones
- Del Examen Físico GeneralDocumento10 páginasDel Examen Físico GeneralJairo Mak SalazarAún no hay calificaciones
- ENDOCRINOLOGÍADocumento2 páginasENDOCRINOLOGÍADanielAún no hay calificaciones
- HORMONASDocumento2 páginasHORMONASDanielAún no hay calificaciones
- D Agcindicesweblibtshb003Documento20 páginasD Agcindicesweblibtshb003DanielAún no hay calificaciones
- Norma Tecnica Programa Nacional en Salud Oral 2018Documento135 páginasNorma Tecnica Programa Nacional en Salud Oral 2018Carlos Eduardo Arze Arana100% (1)
- Aparato Circulatorio - DefinicionDocumento2 páginasAparato Circulatorio - DefinicionDanielAún no hay calificaciones
- Corazón - DEFINICIONDocumento2 páginasCorazón - DEFINICIONDanielAún no hay calificaciones
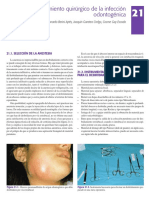
- 21Documento14 páginas21francesca perezAún no hay calificaciones
- Co Ta Ai Historia y Resistencia BacterianaDocumento9 páginasCo Ta Ai Historia y Resistencia BacterianaKaty TaAún no hay calificaciones
- Enfermedades Del Sistema InmunitarioDocumento5 páginasEnfermedades Del Sistema InmunitarioDanielAún no hay calificaciones
- Transtornos HemodinamicosDocumento7 páginasTranstornos HemodinamicosDanielAún no hay calificaciones
- Farmacocinetica 2Documento1 páginaFarmacocinetica 2DanielAún no hay calificaciones
- Dimencionamiento de Equipos MinerosDocumento8 páginasDimencionamiento de Equipos MinerosJuvenal TiconaAún no hay calificaciones
- Recomendaciones para El Uso de EPP COVID 19 PDFDocumento28 páginasRecomendaciones para El Uso de EPP COVID 19 PDFGerlis Tacuri PomaAún no hay calificaciones
- BIOSEGURIDADDocumento13 páginasBIOSEGURIDADDanielAún no hay calificaciones
- Introducción Al Uso de AntibióticosDocumento1 páginaIntroducción Al Uso de AntibióticosDanielAún no hay calificaciones
- Vitamina DDocumento1 páginaVitamina DDanielAún no hay calificaciones
- Fármacos Que Modifican La Homeostasis Del CalcioDocumento1 páginaFármacos Que Modifican La Homeostasis Del CalcioDanielAún no hay calificaciones
- Sistema Estomatognático Morris MizrajiDocumento13 páginasSistema Estomatognático Morris Mizrajipxrxspxlxsxs100% (1)
- Desarrollo de La Denticion PrimariaDocumento31 páginasDesarrollo de La Denticion PrimariaDanielAún no hay calificaciones
- Enfermedades GeneralesDocumento30 páginasEnfermedades GeneralesDanielAún no hay calificaciones
- Vitamina DDocumento1 páginaVitamina DDanielAún no hay calificaciones
- Excreción de FármacosDocumento1 páginaExcreción de FármacosDanielAún no hay calificaciones
- Lesión CelularDocumento8 páginasLesión CelularDanielAún no hay calificaciones
- Crecimiento, Desarrollo y Formación de La Oclusión Capítulo 1 PDFDocumento14 páginasCrecimiento, Desarrollo y Formación de La Oclusión Capítulo 1 PDFJorge ValenzuelaAún no hay calificaciones
- Vih-Sida 1Documento1 páginaVih-Sida 1DanielAún no hay calificaciones
- SIFILISDocumento9 páginasSIFILISDanielAún no hay calificaciones
- Anatomia DentariaDocumento12 páginasAnatomia DentariaDanielAún no hay calificaciones
- Desarrollo PsicosexualDocumento7 páginasDesarrollo PsicosexualDanielAún no hay calificaciones
- Ingenieria de Alimentos - Sharma PDFDocumento358 páginasIngenieria de Alimentos - Sharma PDFAnonymous C06P6LXh9100% (1)
- Manual Operacion AlexionDocumento326 páginasManual Operacion AlexionLILIAN MORALESAún no hay calificaciones
- Uso de La Venda de SmarchDocumento1 páginaUso de La Venda de SmarchMemo50% (2)
- Alumna: Gabriela de Los Ángeles Fonseca Medina. Asignatura: Biología y Conducta. Profesora: Xiomara RodríguezDocumento13 páginasAlumna: Gabriela de Los Ángeles Fonseca Medina. Asignatura: Biología y Conducta. Profesora: Xiomara RodríguezVanessa Guizelly OLVEA ZAPANAAún no hay calificaciones
- Tabla de Calculo Entrega 2Documento37 páginasTabla de Calculo Entrega 2LauraBeltranAún no hay calificaciones
- Harina de Pescado NacionalDocumento1 páginaHarina de Pescado NacionalLaboratorio AgromacaAún no hay calificaciones
- Productos de Hojas de Coca Elaborados en Bolivia Mate de Hojas de CocaDocumento1 páginaProductos de Hojas de Coca Elaborados en Bolivia Mate de Hojas de CocadianaAún no hay calificaciones
- Planes de Beneficios SocialesDocumento4 páginasPlanes de Beneficios SocialesGo TelemedicinaAún no hay calificaciones
- Calificación Test de La FamiliaDocumento4 páginasCalificación Test de La FamiliaDayana Rodriguez100% (1)
- Fundamentacion Teorica Del CuidadoDocumento10 páginasFundamentacion Teorica Del CuidadonatalyAún no hay calificaciones
- Ficha Tecnica Celda SM6Documento2 páginasFicha Tecnica Celda SM6Yeison Steven HernanadezAún no hay calificaciones
- Uso de Repelente NaturalDocumento11 páginasUso de Repelente NaturalGermán DÍAZAún no hay calificaciones
- ATRESIA CONGÉNITA DEL OIDO Orfila2016 PDFDocumento12 páginasATRESIA CONGÉNITA DEL OIDO Orfila2016 PDFEdwin AtencioAún no hay calificaciones
- Melillo, A. & Suarez Ojeda, E. N. (Comp.) - (2001) - Algunos Fundamentos Psicologicos Del Concepto de Resiliencia (83-102) .Documento11 páginasMelillo, A. & Suarez Ojeda, E. N. (Comp.) - (2001) - Algunos Fundamentos Psicologicos Del Concepto de Resiliencia (83-102) .YOAún no hay calificaciones
- Tarea 1 Gestion Humana 1Documento9 páginasTarea 1 Gestion Humana 1Marcos Miguel Sierra FerrerasAún no hay calificaciones
- S33. Litiasis RenalDocumento10 páginasS33. Litiasis RenalMary Tineo BautistaAún no hay calificaciones
- Desatres Naturales Que Fueron Importantes A Nivel Mundial111Documento4 páginasDesatres Naturales Que Fueron Importantes A Nivel Mundial111Jean Pool Puchoc BurgosAún no hay calificaciones
- Conclusiones de Metodos de Medicion de La Calidad Del PescadoDocumento7 páginasConclusiones de Metodos de Medicion de La Calidad Del PescadoFely Silva Ruiz100% (4)
- Fistulas Enterocutaneas Actualidades SobDocumento7 páginasFistulas Enterocutaneas Actualidades SobCecilia SpagnoloAún no hay calificaciones
- Ejemplo Estructura de Guia o Taller de Trabajo de ColegioDocumento2 páginasEjemplo Estructura de Guia o Taller de Trabajo de Colegiocristian cAún no hay calificaciones
- Practica #6. Análisis Granulométricos Por Tamizado.Documento8 páginasPractica #6. Análisis Granulométricos Por Tamizado.Glendys MontielAún no hay calificaciones
- 20 Metodos Analiticos para La Determinacion de Metabolitos Secundarios de PlantasDocumento108 páginas20 Metodos Analiticos para La Determinacion de Metabolitos Secundarios de PlantasAndres SernaAún no hay calificaciones
- La Evolucion de La LactacionDocumento7 páginasLa Evolucion de La LactacionNorma caseresAún no hay calificaciones
- MermeladaDocumento2 páginasMermeladaKelsy NolascoAún no hay calificaciones
- Morbimortalidad NeonatalDocumento50 páginasMorbimortalidad NeonatalJohan BrachoAún no hay calificaciones
- La Biorremediacion Es Una Tecnología Que Aplica Microorganismos para Depurar ContaminantesDocumento6 páginasLa Biorremediacion Es Una Tecnología Que Aplica Microorganismos para Depurar ContaminantesMatias CandelaAún no hay calificaciones
- REINO: Plantae Division: Subdivisión: Angiospermae Clase: Orden: Familia: Genero: EspecieDocumento19 páginasREINO: Plantae Division: Subdivisión: Angiospermae Clase: Orden: Familia: Genero: EspecierAún no hay calificaciones
- Ìká ÒfúnDocumento8 páginasÌká ÒfúnJuanci Rodriguez100% (1)
- Superacion de TraumasDocumento1 páginaSuperacion de TraumasBerly Tovar VergaraAún no hay calificaciones
- Que Es Cpi y CPI en Cuantificación Por Factores en Cromatografía de Factor InternoDocumento21 páginasQue Es Cpi y CPI en Cuantificación Por Factores en Cromatografía de Factor InternoEnauris MateoAún no hay calificaciones