También podría gustarte
- Los inicios de la vida: La evolución en la Tierra precámbricaDe EverandLos inicios de la vida: La evolución en la Tierra precámbricaAún no hay calificaciones
- Laboratorio No.3Documento3 páginasLaboratorio No.3eliza garciaAún no hay calificaciones
- Evolución - Lectura 2Documento10 páginasEvolución - Lectura 2Alejandro SaavedraAún no hay calificaciones
- Definición y Concepto de BiologíaDocumento19 páginasDefinición y Concepto de BiologíaAna SandovalAún no hay calificaciones
- Teorias EvolutivasDocumento5 páginasTeorias EvolutivasAmanda FloresAún no hay calificaciones
- Los Hitos Del Conocimiento BiológicoDocumento5 páginasLos Hitos Del Conocimiento BiológicoJuan Manuel Castro Pérez100% (1)
- Resumen de Dos Articuos de EvolucionDocumento4 páginasResumen de Dos Articuos de EvolucionADELFO MORALES GONZALEZAún no hay calificaciones
- BotanicaDocumento41 páginasBotanicaMauricio Giraldo DiazAún no hay calificaciones
- La Teoría de La SimbiogénesisDocumento5 páginasLa Teoría de La SimbiogénesisEiner Sepúlveda Zúñiga100% (1)
- Que Es La Epigenetica What Is EpigenetiDocumento25 páginasQue Es La Epigenetica What Is EpigenetiEmiliano Sevilleja MartinezAún no hay calificaciones
- Desarrollo Historico de La Genetica de PoblacionesDocumento3 páginasDesarrollo Historico de La Genetica de Poblacionesenith johanaAún no hay calificaciones
- BiologÍa Evolutiva Del DesarrolloDocumento9 páginasBiologÍa Evolutiva Del Desarrollomarilynday12Aún no hay calificaciones
- Teorías de La Evolución ESFMDocumento9 páginasTeorías de La Evolución ESFMYesenia VitoAún no hay calificaciones
- Heterocronía en AnurosDocumento20 páginasHeterocronía en AnurosJavier RojasAún no hay calificaciones
- Teoría de La Evolución Aristoteles, Darwin, LamarckDocumento18 páginasTeoría de La Evolución Aristoteles, Darwin, LamarckKarla Osorio67% (3)
- BIOLOGIADocumento6 páginasBIOLOGIAholasoycitla27Aún no hay calificaciones
- Biología CelularDocumento12 páginasBiología CelularLmt Luis ReyesAún no hay calificaciones
- Evo DevoDocumento4 páginasEvo DevoMer OyhAún no hay calificaciones
- Investigación de BiologíaDocumento4 páginasInvestigación de BiologíaAlexander RuizAún no hay calificaciones
- Síntesis Evolutiva ModernaDocumento4 páginasSíntesis Evolutiva ModernaLorena Gámez AlcaláAún no hay calificaciones
- Biologia Conceptos BasicosDocumento41 páginasBiologia Conceptos BasicosSolitario IncomprendidoAún no hay calificaciones
- Síntesis Evolutiva ModernaDocumento27 páginasSíntesis Evolutiva ModernajoanAún no hay calificaciones
- Theodosius DobzhanskyDocumento4 páginasTheodosius Dobzhanskysad_jrAún no hay calificaciones
- Situación Actual y Perspectivas de La BotánicaDocumento5 páginasSituación Actual y Perspectivas de La BotánicaEnmanuel GilAún no hay calificaciones
- Dialnet GeneYMutacion 62135Documento20 páginasDialnet GeneYMutacion 62135LenardR.Apaza100% (1)
- Margulis PDFDocumento1 páginaMargulis PDFCarla Vidal ValdebenitoAún no hay calificaciones
- FilogeniaDocumento3 páginasFilogeniaJared KortAún no hay calificaciones
- MurcielagoDocumento5 páginasMurcielagoAbiertamentePnlCali TalleresAún no hay calificaciones
- Mor Fo GenesisDocumento10 páginasMor Fo GenesiszhikitaloAún no hay calificaciones
- La Síntesis Moderna en Biología - PedrocheDocumento7 páginasLa Síntesis Moderna en Biología - PedrocheJuan Antonio Valls FerrerAún no hay calificaciones
- Stephen Meyer El Origen de La Informacion BiologicaDocumento30 páginasStephen Meyer El Origen de La Informacion BiologicadeimosyfobosAún no hay calificaciones
- Evolución y Sus ClasificacionesDocumento6 páginasEvolución y Sus Clasificacionesjessfey ggAún no hay calificaciones
- El Lenguaje de La NaturalezaDocumento33 páginasEl Lenguaje de La NaturalezaDaniel HidalgoAún no hay calificaciones
- Naturales Biologia - Guia EvolucionDocumento6 páginasNaturales Biologia - Guia EvolucionauraAún no hay calificaciones
- Teoria Sintetica de La Evolucion2Documento6 páginasTeoria Sintetica de La Evolucion2Nahomi SantillanaAún no hay calificaciones
- N 7 Teoría Sintética EvolutivaDocumento13 páginasN 7 Teoría Sintética EvolutivaRafaela YanezAún no hay calificaciones
- Artículo de Revista - Simbiosis y Evolución - Nicolás José Lavagnino, Alicia Massarini y Guillermo FolgueraDocumento21 páginasArtículo de Revista - Simbiosis y Evolución - Nicolás José Lavagnino, Alicia Massarini y Guillermo FolgueraFranco VillalbaAún no hay calificaciones
- A4 Portada de Propuesta de Proyecto Científico Infantil Doodle Azul y NaranjaDocumento8 páginasA4 Portada de Propuesta de Proyecto Científico Infantil Doodle Azul y NaranjaMya QueenAún no hay calificaciones
- FilogeniaDocumento7 páginasFilogeniaiOSvaldo 7Aún no hay calificaciones
- Los Naturalistas Charles Darwin y Alfred Russel WallaceDocumento5 páginasLos Naturalistas Charles Darwin y Alfred Russel Wallacefrancisco islasAún no hay calificaciones
- Capitulo-9 Filogenia Evolucion MolecularDocumento29 páginasCapitulo-9 Filogenia Evolucion Molecularmarlynfg27Aún no hay calificaciones
- Evo Devo Hacia Un Nuevo Paradigma en Biolog A EvolutivaDocumento3 páginasEvo Devo Hacia Un Nuevo Paradigma en Biolog A EvolutivamagiqoAún no hay calificaciones
- Filosofia de La BiologiaDocumento15 páginasFilosofia de La BiologiaJaime Andres Vasquez MarchantAún no hay calificaciones
- Antonio Lazcano Araujo - El Ultimo Ancestro Común PDFDocumento9 páginasAntonio Lazcano Araujo - El Ultimo Ancestro Común PDFgonzalezpuertoAún no hay calificaciones
- ReseñaDocumento7 páginasReseñaIngrid AcatlAún no hay calificaciones
- Evolución en MéxicoDocumento9 páginasEvolución en MéxicoDeth GonzálezAún no hay calificaciones
- El Origen de La Información Biológica y Las Categorías Taxonómicas Superiores PDFDocumento30 páginasEl Origen de La Información Biológica y Las Categorías Taxonómicas Superiores PDFVitelioAún no hay calificaciones
- ANTROPOSMODERNO El Lenguaje de La NaturalezaDocumento23 páginasANTROPOSMODERNO El Lenguaje de La NaturalezaAntonio Francisco MuñozAún no hay calificaciones
- A6 P2 Teroria Sintetica BLG NJCMDocumento3 páginasA6 P2 Teroria Sintetica BLG NJCMNayma Julie Contreras MolinaAún no hay calificaciones
- Ensayo Microbiologia Ideas AbsurdasDocumento2 páginasEnsayo Microbiologia Ideas AbsurdasAnderson David Bolano BedoyaAún no hay calificaciones
- Filogenia y Evolucion Molecular-RevistaDocumento30 páginasFilogenia y Evolucion Molecular-RevistaPier Jhoani Ñañez LlanosAún no hay calificaciones
- EvolucionDocumento10 páginasEvolucionMartinez Marinez ClarissaAún no hay calificaciones
- De Asúa-EvoluciónDocumento22 páginasDe Asúa-EvoluciónMarian MagAún no hay calificaciones
- Introducción A La BiologíaDocumento31 páginasIntroducción A La BiologíaAnallely López0% (1)
- Ensayo 1.despliegue - de - La - VidaDocumento3 páginasEnsayo 1.despliegue - de - La - VidaAlisson Camacho ErazoAún no hay calificaciones
- Hormonas: Mensajeros químicos y comunicación celularDe EverandHormonas: Mensajeros químicos y comunicación celularCalificación: 2.5 de 5 estrellas2.5/5 (2)
- Mitología EgipciaDocumento23 páginasMitología EgipciaAlejandro Morante MedranoAún no hay calificaciones
- Mordeduras SerpientesDocumento4 páginasMordeduras Serpientesscribdraul123Aún no hay calificaciones
- NormalidadDocumento7 páginasNormalidadOscar Javier Salazar MezaAún no hay calificaciones
- El Origen de Los MamiferosDocumento34 páginasEl Origen de Los MamiferosOrlando BacaAún no hay calificaciones
- 1080073248Documento60 páginas1080073248Oscar Javier Salazar MezaAún no hay calificaciones
- 2 SauveDocumento30 páginas2 SauveDiana Laura PerezAún no hay calificaciones
- Espiral 1665-0565: Issn: Espiral@fuentes - Csh.udg - MXDocumento32 páginasEspiral 1665-0565: Issn: Espiral@fuentes - Csh.udg - MXOscar Javier Salazar MezaAún no hay calificaciones
- Proyecto. Vulnerabilidad ClimáticaDocumento4 páginasProyecto. Vulnerabilidad ClimáticaOscar Javier Salazar MezaAún no hay calificaciones
- Construcción de Escalas de Actitud Tipo Thurst y LikertDocumento50 páginasConstrucción de Escalas de Actitud Tipo Thurst y Likertnegro_pinolero9621Aún no hay calificaciones
- AAVV - Evolución - La Base de La BiologíaDocumento535 páginasAAVV - Evolución - La Base de La BiologíaPaul Dante Carranco Blanquet100% (2)
- El Sistema Producto CaféDocumento486 páginasEl Sistema Producto CaféOscar Javier Salazar Meza92% (12)
- Plaguicidas AgricolasDocumento17 páginasPlaguicidas AgricolasFITOTECNIAJUAREZAún no hay calificaciones
- Plaguicidas AgricolasDocumento17 páginasPlaguicidas AgricolasFITOTECNIAJUAREZAún no hay calificaciones
- Resi DuosDocumento7 páginasResi DuosOscar Javier Salazar MezaAún no hay calificaciones
- 3 PlatelmintosDocumento34 páginas3 PlatelmintosOscar Javier Salazar MezaAún no hay calificaciones
- Phyllum Artropoda Practica 1Documento5 páginasPhyllum Artropoda Practica 1Oscar Javier Salazar MezaAún no hay calificaciones
- TrilobitesDocumento6 páginasTrilobitesOscar Javier Salazar MezaAún no hay calificaciones
- Trictico - Seguridad en La Operacion Maquinaria A.Documento2 páginasTrictico - Seguridad en La Operacion Maquinaria A.Alejandra Gomez100% (1)
- Taller Convertidores CCaCCDocumento6 páginasTaller Convertidores CCaCCDavidAcevedoTrejosAún no hay calificaciones
- RP Cta5 k14 FichaDocumento20 páginasRP Cta5 k14 Ficharosa luz100% (1)
- Departamento de UcayaliDocumento6 páginasDepartamento de UcayaliDana Dxr'kAún no hay calificaciones
- Taller Sobre Estudios Epidemiológicos y Medidas de Riesgo Ejercicio 1Documento2 páginasTaller Sobre Estudios Epidemiológicos y Medidas de Riesgo Ejercicio 1estefania loperaAún no hay calificaciones
- Formato de Inspeccion de EslingasDocumento6 páginasFormato de Inspeccion de Eslingasronald niño0% (1)
- Bases Contratacion Directa AlimentosDocumento55 páginasBases Contratacion Directa AlimentosJoiss ChirinosAún no hay calificaciones
- INFORME DE RESUSPENSION DE MATERIA PARTICULADO ModificadoDocumento16 páginasINFORME DE RESUSPENSION DE MATERIA PARTICULADO ModificadoElvis HernandezAún no hay calificaciones
- Tarea V de Gastronomía DominicanaDocumento12 páginasTarea V de Gastronomía DominicanaErmil Manuel Adames GómezAún no hay calificaciones
- Bases Físicas de La EcografíaDocumento79 páginasBases Físicas de La Ecografíaterebgarcia100% (2)
- 1409 0015 MLCR 33 01 00063 PDFDocumento7 páginas1409 0015 MLCR 33 01 00063 PDFRodrigo José BustamanteAún no hay calificaciones
- Formatos de Fichas Textual y de ResumenDocumento3 páginasFormatos de Fichas Textual y de ResumenAVILA CÁCEDA Ingrid ElenaAún no hay calificaciones
- Antamina DiapoDocumento18 páginasAntamina DiapoLorena Nunura AgüeroAún no hay calificaciones
- PDFDocumento669 páginasPDFfabri1909Aún no hay calificaciones
- Mecánica de Fluidos - Usmp 3, HidrostaticaDocumento11 páginasMecánica de Fluidos - Usmp 3, HidrostaticaPercy AJAún no hay calificaciones
- Salvado de MaizDocumento2 páginasSalvado de Maizigelfritz5407Aún no hay calificaciones
- Modelos Con Sus SolucionesDocumento49 páginasModelos Con Sus SolucionesJuan Pablo Requez100% (2)
- Informe Final Hormigones Especiales Tecnologia Del HormigonDocumento55 páginasInforme Final Hormigones Especiales Tecnologia Del HormigonFrancisco Martinez ParraguezAún no hay calificaciones
- Acta 03 de 2009 Sobre SaboresDocumento5 páginasActa 03 de 2009 Sobre SaborescmgabrieladezAún no hay calificaciones
- Avermectina FarmacologiaDocumento16 páginasAvermectina FarmacologiaVivi GhiringhelliAún no hay calificaciones
- Ejercicio 28 Estadistica Descriptiva222Documento9 páginasEjercicio 28 Estadistica Descriptiva222Fabian Andres Garcia Perez100% (4)
- Actividad 8 Actividad Evaluativa Investigacion de EventosDocumento29 páginasActividad 8 Actividad Evaluativa Investigacion de EventosLuis Miguel MoralesAún no hay calificaciones
- Desarrollo Actividad 2 PasteleriaDocumento4 páginasDesarrollo Actividad 2 PasteleriaCarlos Adonnai CardenasAún no hay calificaciones
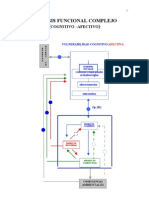
- Walter Riso - Análisis Funcional ComplejoDocumento8 páginasWalter Riso - Análisis Funcional ComplejoSamantha Rojas GuzmanAún no hay calificaciones
- A Mover El Esqueleto - Transformarse TransformandoDocumento9 páginasA Mover El Esqueleto - Transformarse TransformandoManu CarrascoAún no hay calificaciones
- Manual de Practicas de Laboratorio Del Sistema Nervioso Q1-2015Documento65 páginasManual de Practicas de Laboratorio Del Sistema Nervioso Q1-2015RogelioEucedaAún no hay calificaciones
- La Célula y Sus Antecedentes HistóricosDocumento3 páginasLa Célula y Sus Antecedentes HistóricosElizabeth OrtegaAún no hay calificaciones
- Cargador de Batería Con Fuente PC ATXDocumento10 páginasCargador de Batería Con Fuente PC ATXFelix Ruiz100% (1)
- XYPEX - APLICACION SISTEMA ImpermeabilizanteDocumento8 páginasXYPEX - APLICACION SISTEMA Impermeabilizantechristian paul santur cunyaracheAún no hay calificaciones
- Evaluación Y Tratamiento Psicológico de Un Caso de Cáncer de Colon Con Fobia A La Quimioterapia Y Trastorno AdaptativoDocumento11 páginasEvaluación Y Tratamiento Psicológico de Un Caso de Cáncer de Colon Con Fobia A La Quimioterapia Y Trastorno AdaptativoConstanza CuevasAún no hay calificaciones