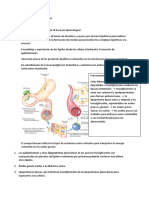
También podría gustarte
- Examen Parcial de Bioquimica 2Documento8 páginasExamen Parcial de Bioquimica 2darlin60% (5)
- Anatomía de La OrbitaDocumento12 páginasAnatomía de La OrbitaSofia Leiva JaraAún no hay calificaciones
- Clase 1 Orbita y Anexos OcularesDocumento56 páginasClase 1 Orbita y Anexos OcularesSofia Leiva Jara67% (3)
- Disfunción Mitocondrial y Enfermedades CardiacasDocumento15 páginasDisfunción Mitocondrial y Enfermedades CardiacasdrafecanixthAún no hay calificaciones
- El Magnifico MagnesioDocumento496 páginasEl Magnifico MagnesioPablo Mestre Almendras100% (1)
- Topografia Corneal 2 ParteDocumento39 páginasTopografia Corneal 2 ParteSofia Leiva JaraAún no hay calificaciones
- Test MetabolismoDocumento27 páginasTest MetabolismoDaniel Moreno GarciaAún no hay calificaciones
- Manual de Neurooftalmologia PDFDocumento170 páginasManual de Neurooftalmologia PDFSofia Leiva JaraAún no hay calificaciones
- 2017 Guia RetinosDocumento7 páginas2017 Guia RetinosSofia Leiva JaraAún no hay calificaciones
- Javier BUCETA FERNÁNDEZ - Elka Radoslavova KOROUTCHEVA - Juan Manuel PASTOR RUIZ - Temas de Biofísica (2012, Editorial UNED)Documento340 páginasJavier BUCETA FERNÁNDEZ - Elka Radoslavova KOROUTCHEVA - Juan Manuel PASTOR RUIZ - Temas de Biofísica (2012, Editorial UNED)Andrés Lopez100% (4)
- CS FinalDocumento9 páginasCS FinalSofia Leiva JaraAún no hay calificaciones
- Angiografía Análoga TDDocumento8 páginasAngiografía Análoga TDSofia Leiva JaraAún no hay calificaciones
- Manual de Exámenes en AdultosDocumento7 páginasManual de Exámenes en AdultosSofia Leiva JaraAún no hay calificaciones
- Músculos Del DorsoDocumento8 páginasMúsculos Del DorsoSofia Leiva JaraAún no hay calificaciones
- Capitulo 04 Trabajo y Energia - Item de TestDocumento10 páginasCapitulo 04 Trabajo y Energia - Item de TestSofia Leiva JaraAún no hay calificaciones
- Clase 2Documento9 páginasClase 2Sofia Leiva JaraAún no hay calificaciones
- Capacitores TeoriaDocumento12 páginasCapacitores TeoriaDaniel Garcia PinosAún no hay calificaciones
- Tema 4 Metabolismo y Procesos EnergéticosDocumento58 páginasTema 4 Metabolismo y Procesos EnergéticosDeilianny HernandezAún no hay calificaciones
- El Ciclo de KrebsDocumento18 páginasEl Ciclo de KrebsErick EspinozaAún no hay calificaciones
- Propiedades, Beneficios y Efectos de La Guanábana (Annona Muricata L.) Sobre La Glucemia y El CáncerDocumento16 páginasPropiedades, Beneficios y Efectos de La Guanábana (Annona Muricata L.) Sobre La Glucemia y El CáncerJhenny ArancibiaAún no hay calificaciones
- "Bioenergética:": Libera ExotérmicaDocumento21 páginas"Bioenergética:": Libera ExotérmicaMARÍA BARROSAún no hay calificaciones
- Clase Fosforilacion OxidativaDocumento19 páginasClase Fosforilacion OxidativavalentinaAún no hay calificaciones
- Tema 12 Metabolismo de Lípidos 2021-1Documento160 páginasTema 12 Metabolismo de Lípidos 2021-1Gustavo Alonzo Samame GoicocheaAún no hay calificaciones
- Metabolismo de Los LípidosDocumento2 páginasMetabolismo de Los LípidosJaviera Gotuzzo CarrascoAún no hay calificaciones
- Cuestionario MetabolismoDocumento3 páginasCuestionario MetabolismoIsamar QuispeAún no hay calificaciones
- Glucólisis y GluconeogenesisDocumento2 páginasGlucólisis y Gluconeogenesischula98Aún no hay calificaciones
- Manejo de La Hipoxia FisiopatologíaDocumento14 páginasManejo de La Hipoxia FisiopatologíaDavid MamaniAún no hay calificaciones
- Taller BiomoleculasDocumento21 páginasTaller BiomoleculasJuliana VelásquezAún no hay calificaciones
- 1F - 3T - Metabolismo Via GlicolíticaDocumento42 páginas1F - 3T - Metabolismo Via GlicolíticaLidiaAlejandraRiveraPonceAún no hay calificaciones
- Microb - RetroalimentaciónDocumento289 páginasMicrob - RetroalimentaciónViolet' Armas ChavezAún no hay calificaciones
- Catabolismo y AnabolismoDocumento2 páginasCatabolismo y AnabolismoRamiro Amari100% (1)
- TALLER DE CONCEPTOS DE METABOLISMO Y BIOENERGÉTICA. Biología 2021Documento4 páginasTALLER DE CONCEPTOS DE METABOLISMO Y BIOENERGÉTICA. Biología 2021Alex NarvaezAún no hay calificaciones
- Preguntas Bio COMPLETODocumento36 páginasPreguntas Bio COMPLETOsabrina100% (1)
- Guia No 8 BioenergeticaDocumento6 páginasGuia No 8 Bioenergeticade_jvAún no hay calificaciones
- Lipidos 1Documento24 páginasLipidos 1Generación para DIOSAún no hay calificaciones
- Alteracion de La Actividad EnzimaticaDocumento11 páginasAlteracion de La Actividad EnzimaticaFatima AlvaradoAún no hay calificaciones
- Los CarbohidratosDocumento22 páginasLos CarbohidratosGilberto Gutierrez RivasAún no hay calificaciones
- Cadena Respiratoria Grado 10 - 3Documento8 páginasCadena Respiratoria Grado 10 - 3Mariana RuizAún no hay calificaciones
- Libritos Negros C.B.C.Documento123 páginasLibritos Negros C.B.C.Pablo SánchezAún no hay calificaciones
- Alimentacion de La Vaca LecheraDocumento3 páginasAlimentacion de La Vaca LecheraEnriq Sisson IIIAún no hay calificaciones
- ANTIOXIDANTESDocumento3 páginasANTIOXIDANTESAlex FarkAún no hay calificaciones
- INFOGRAFIADocumento1 páginaINFOGRAFIAYami YamAún no hay calificaciones