También podría gustarte
- Endotermia y Ectotermia Como Mecanismos de Termorregulacion enDocumento10 páginasEndotermia y Ectotermia Como Mecanismos de Termorregulacion endiecolo26Aún no hay calificaciones
- Reflejos Por Nivel MaduraciónDocumento3 páginasReflejos Por Nivel MaduraciónTatiana Ordoñez100% (1)
- Introducción: Revista Colombiana de Ciencia Animal, Vol. 4, No. 1, 2011Documento6 páginasIntroducción: Revista Colombiana de Ciencia Animal, Vol. 4, No. 1, 2011Rebeca GuerreroAún no hay calificaciones
- Tarea 6 Morfofisiologia AnimalDocumento5 páginasTarea 6 Morfofisiologia Animalgonzalo hernandez100% (1)
- Mecanismo de TermoregulacionDocumento5 páginasMecanismo de TermoregulacionAnonymous S0BZX8Aún no hay calificaciones
- Trabajo de TermorregulacionDocumento23 páginasTrabajo de Termorregulaciondanna quintanaAún no hay calificaciones
- Unidad Ii y IiiDocumento18 páginasUnidad Ii y IiiGladys NoemÕ Cauich KÏAún no hay calificaciones
- Regulación de La Temperatura CorporalDocumento5 páginasRegulación de La Temperatura CorporalyesseniaAún no hay calificaciones
- Unidad V Calor y FríoDocumento21 páginasUnidad V Calor y FríoZzheiLa'h OteRo'hAún no hay calificaciones
- EcofisiologiaDocumento8 páginasEcofisiologiaAngie DiazAún no hay calificaciones
- Practica 17 TermoregulacionDocumento9 páginasPractica 17 TermoregulacionAitana PiñaAún no hay calificaciones
- La Vida Animal Es Posible Debido A Los Sistemas de Adaptación Térmica Tanto A Los Extremos de Temperaturas Antes Señalados Como A La Variedad de Rangos Climáticos en Las Diferentes Áreas Del PlanetaDocumento3 páginasLa Vida Animal Es Posible Debido A Los Sistemas de Adaptación Térmica Tanto A Los Extremos de Temperaturas Antes Señalados Como A La Variedad de Rangos Climáticos en Las Diferentes Áreas Del Planetakevin granadosAún no hay calificaciones
- TERMORREGULACIÓNDocumento9 páginasTERMORREGULACIÓNpaolaAún no hay calificaciones
- TermorregulaciónDocumento5 páginasTermorregulaciónAlejandra CallejasAún no hay calificaciones
- Regulación Térmica I y IIDocumento16 páginasRegulación Térmica I y IICleyver RodriguezAún no hay calificaciones
- Zona de Confort para VacunosDocumento20 páginasZona de Confort para VacunosPablo VasquezAún no hay calificaciones
- Regulacón y Homestasis de La Temperatrura..Documento6 páginasRegulacón y Homestasis de La Temperatrura..yojaxci alexandra mora paezAún no hay calificaciones
- Morfofisiología Animal-1Documento7 páginasMorfofisiología Animal-1juan daniel florez diazAún no hay calificaciones
- Fisiología de La TermoregulaciónDocumento4 páginasFisiología de La TermoregulaciónMARIA LUISA ALZATE RINCONAún no hay calificaciones
- TermoregulaciónDocumento10 páginasTermoregulaciónClara Elena VeraAún no hay calificaciones
- Mantenimiento CorporalDocumento15 páginasMantenimiento CorporalMiguel CabreraAún no hay calificaciones
- Guia TermorregulacionDocumento15 páginasGuia TermorregulacionAllison OportaAún no hay calificaciones
- Temperatura Corporal en Animales Grupo 11Documento8 páginasTemperatura Corporal en Animales Grupo 11María HernándezAún no hay calificaciones
- Animales HomeotermosDocumento20 páginasAnimales HomeotermosRicardo Martos AcuñaAún no hay calificaciones
- Fisiología Reporte IIDocumento16 páginasFisiología Reporte IIChristopher VelázquezAún no hay calificaciones
- Trabajo Sobre Regulacion de La Temperatura PDFDocumento10 páginasTrabajo Sobre Regulacion de La Temperatura PDFGabriel FuentesAún no hay calificaciones
- Zona de Confort para VacunosDocumento5 páginasZona de Confort para VacunosPablo VasquezAún no hay calificaciones
- Sistema Regulador de La Temperatur 3Documento12 páginasSistema Regulador de La Temperatur 3Angelys Del Valle Gómez SalazarAún no hay calificaciones
- TERMOREGUALCIONDocumento5 páginasTERMOREGUALCIONꓟᴄRḼṎṢ ẎẺịṆṦyAún no hay calificaciones
- Homeostasis Termo y OsmoDocumento76 páginasHomeostasis Termo y OsmoAna Karina Tarco Perez0% (1)
- Regulacion de La Temperatura CorporalDocumento4 páginasRegulacion de La Temperatura CorporalOscar VejarAún no hay calificaciones
- Articulo de Termoregulacion 23 de Mayo 2019Documento10 páginasArticulo de Termoregulacion 23 de Mayo 2019rocioAún no hay calificaciones
- Mecanismos de HomeostasisDocumento7 páginasMecanismos de HomeostasisdrcidbaezAún no hay calificaciones
- Homeostasis Termo y OsmoDocumento76 páginasHomeostasis Termo y OsmocalleleoAún no hay calificaciones
- La Termorregulación en AnimalesDocumento12 páginasLa Termorregulación en AnimalesvalentinaAún no hay calificaciones
- Homeostasis Termorregulacion Y OsmorregulacionDocumento76 páginasHomeostasis Termorregulacion Y OsmorregulacionmeteorizadaAún no hay calificaciones
- HomeostasisDocumento6 páginasHomeostasisACUARIO COMPUTERSAún no hay calificaciones
- FCOM - Regulacion de La Temperatura CorporalDocumento7 páginasFCOM - Regulacion de La Temperatura CorporalJoshep InAún no hay calificaciones
- Tarea 4 - Ecofisiología y Su Importancia en Producción Animal 1Documento12 páginasTarea 4 - Ecofisiología y Su Importancia en Producción Animal 1Haiberth DoradoAún no hay calificaciones
- TermorregulacionDocumento17 páginasTermorregulacionNiza Jarintzi Sebastian VillaAún no hay calificaciones
- 3714.pdf# - Text Existen Mecanismos de Regulación Que, y La Evaporación (12) .Documento24 páginas3714.pdf# - Text Existen Mecanismos de Regulación Que, y La Evaporación (12) .Guadalupe MartinezAún no hay calificaciones
- Efecto de La Temperatura Ambiental y La Humedad Relativa Sobre La Temperatura Corporal en Caprinos PDFDocumento10 páginasEfecto de La Temperatura Ambiental y La Humedad Relativa Sobre La Temperatura Corporal en Caprinos PDFJuanDiegoNarvaezBastidasAún no hay calificaciones
- Mecanismos Fisiológicos de La Termorregulación en ReptilesDocumento12 páginasMecanismos Fisiológicos de La Termorregulación en ReptilesSadiq NayrufAún no hay calificaciones
- ECOLOGIADocumento3 páginasECOLOGIAKiara IbañezAún no hay calificaciones
- Homeostasis - BiologiaDocumento5 páginasHomeostasis - BiologiaNagbys Obando de HernándezAún no hay calificaciones
- Biofisica Clase 13 2018 II Practica Termodinamica Practica N 12Documento4 páginasBiofisica Clase 13 2018 II Practica Termodinamica Practica N 12Wildor CastilloAún no hay calificaciones
- Unidad 3 - Tarea 4 - EcofisiologíaDocumento7 páginasUnidad 3 - Tarea 4 - Ecofisiologíaeduardo arenas67% (3)
- Guion TermoregulacionDocumento3 páginasGuion TermoregulacionDIANA LIZETTE ROJAS GARCIAAún no hay calificaciones
- HipotermiaDocumento4 páginasHipotermiaJóse santiago Díaz giraldoAún no hay calificaciones
- Taller TermorregulacionDocumento5 páginasTaller TermorregulacionIsadora HernándezAún no hay calificaciones
- EnsayoDocumento8 páginasEnsayoAlejandro CardozoAún no hay calificaciones
- TERMOREGULACIONDocumento11 páginasTERMOREGULACIONAnaly De LeonAún no hay calificaciones
- Transcripciones Dr. CuadraDocumento243 páginasTranscripciones Dr. CuadraElias Emmanuel JaimeAún no hay calificaciones
- HomeotermosDocumento9 páginasHomeotermosVilca Mescco DicksonAún no hay calificaciones
- Mecanismos Fisicos y Fisiologicos de Adaptacion de Los Animales HomeotermosDocumento19 páginasMecanismos Fisicos y Fisiologicos de Adaptacion de Los Animales HomeotermosIDELSOAún no hay calificaciones
- C22 - Nutrición Deportiva 2Documento54 páginasC22 - Nutrición Deportiva 2Lily-Gira MolinaAún no hay calificaciones
- TermorregulaciónDocumento7 páginasTermorregulaciónoscarAún no hay calificaciones
- TERMORREGULACION Equipo 7Documento29 páginasTERMORREGULACION Equipo 7Andrea HernandezAún no hay calificaciones
- Termorregulacion, Hipertermia, HipotermiaDocumento24 páginasTermorregulacion, Hipertermia, HipotermiaSergio ReyesAún no hay calificaciones
- El cambio climático y la biología funcional de los organismosDe EverandEl cambio climático y la biología funcional de los organismosAún no hay calificaciones
- Plantas Que Curan Enfermedades Segun La Enseñanza RosacruzDe EverandPlantas Que Curan Enfermedades Segun La Enseñanza RosacruzAún no hay calificaciones
- Clasificacion de Las Neoplasias en Los Animales DomesticosDocumento26 páginasClasificacion de Las Neoplasias en Los Animales DomesticosJ.M.EAún no hay calificaciones
- Cartilago y Hueso-GartnerDocumento25 páginasCartilago y Hueso-Gartnerpablitss27100% (1)
- Apicultura en Colombia PDFDocumento16 páginasApicultura en Colombia PDFFaval CastroAún no hay calificaciones
- Tabla NutricionalDocumento1 páginaTabla NutricionalJ.M.EAún no hay calificaciones
- Aponeurosis de La Region Antero Lateral Del ToraxDocumento3 páginasAponeurosis de La Region Antero Lateral Del ToraxJorge Cornelio Mainato Shagñay0% (1)
- Tumores Sólidos PancreaticosDocumento18 páginasTumores Sólidos PancreaticosGiancarlo MorenoAún no hay calificaciones
- Hemato 1er ParcialDocumento110 páginasHemato 1er ParcialRodrigo Caballero DíazAún no hay calificaciones
- PLEURADocumento224 páginasPLEURAGustavo CarraroAún no hay calificaciones
- Der Rame Cerebral 1Documento13 páginasDer Rame Cerebral 1Lili MaciasAún no hay calificaciones
- Xi ChuanDocumento56 páginasXi ChuanFloFlóAún no hay calificaciones
- Cuadro de Examen Fisico y Transc.a La HCDocumento9 páginasCuadro de Examen Fisico y Transc.a La HCMicTrujilloCapAún no hay calificaciones
- Irrigación de La Glándula TiroidesDocumento4 páginasIrrigación de La Glándula TiroidesNicole Antonella RiosAún no hay calificaciones
- ULTRASONIDODocumento14 páginasULTRASONIDOChristian AugustoAún no hay calificaciones
- Irrigacion UterinaDocumento7 páginasIrrigacion UterinaPaolita León Del CarpioAún no hay calificaciones
- ¿Gripe o Zika?Documento16 páginas¿Gripe o Zika?msanchezpi100% (1)
- Aquí Hay Más de 2000 Refranes TradicionalesDocumento54 páginasAquí Hay Más de 2000 Refranes TradicionalesBeto Luna PerezAún no hay calificaciones
- Panthera Onca (Linnaeus, 1758)Documento11 páginasPanthera Onca (Linnaeus, 1758)eliizacndAún no hay calificaciones
- DermatitisDocumento8 páginasDermatitisSaulo Salomon Mija ReynaAún no hay calificaciones
- GuanacoDocumento6 páginasGuanacoLore CuadraAún no hay calificaciones
- Infecciones VaginalesDocumento26 páginasInfecciones VaginalesSanchez2326100% (1)
- Teoria de Los Elementos y Bases de La AcupunturaDocumento34 páginasTeoria de Los Elementos y Bases de La Acupunturanathalie fortuny100% (1)
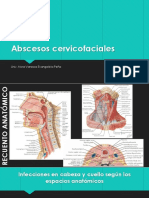
- Abscesos CervicofacialesDocumento17 páginasAbscesos CervicofacialesJota Orellana CuellarAún no hay calificaciones
- Mnemotecnia Pediatria PDFDocumento1 páginaMnemotecnia Pediatria PDFzramon1006Aún no hay calificaciones
- Clasificación de Los MicroorganismosDocumento34 páginasClasificación de Los MicroorganismosRodrigo Pinto100% (1)
- Obtencion de ColesterolDocumento5 páginasObtencion de ColesterolJaime SanAún no hay calificaciones
- PleurasDocumento18 páginasPleurasJavier VargasAún no hay calificaciones
- Comidas Tipicas y Revetas en INGLES Y CASTELLANODocumento19 páginasComidas Tipicas y Revetas en INGLES Y CASTELLANOMamacita Bella de EstradaAún no hay calificaciones
- Physio Ex Ejercicio 9 Actividad 1fisiologia RenalDocumento4 páginasPhysio Ex Ejercicio 9 Actividad 1fisiologia RenalLucia Saavedra LozadaAún no hay calificaciones
- Stormy Glenn - Mi ChicaDocumento135 páginasStormy Glenn - Mi ChicaMariannyAlexandraChirinos67% (3)
- JJMillás - de Corpore Insepulto (Poesia)Documento2 páginasJJMillás - de Corpore Insepulto (Poesia)My SelfAún no hay calificaciones
- Dioses Relacionados Con La MedicinaDocumento6 páginasDioses Relacionados Con La MedicinaInod MorenoAún no hay calificaciones
- Información Sobre El LoboDocumento16 páginasInformación Sobre El Lobovannezzca100% (1)
- Tratamiento de La Diarrea Del ViajeroDocumento2 páginasTratamiento de La Diarrea Del ViajeroKarol Michelle Bravo BorjasAún no hay calificaciones