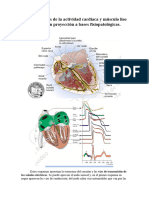
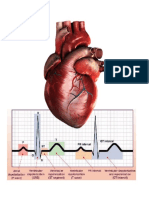
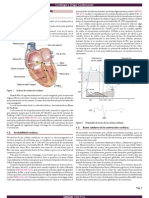
También podría gustarte
- Enero-Febrero 2023 Poda en El CerebroDocumento91 páginasEnero-Febrero 2023 Poda en El CerebroLucia Del Sol100% (4)
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)De EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Calificación: 5 de 5 estrellas5/5 (9)
- Harrison Principios de Medicina Interna - 20º - Tomo 2 - Unlocked (PDF - Io)Documento12 páginasHarrison Principios de Medicina Interna - 20º - Tomo 2 - Unlocked (PDF - Io)Cristina Lissette PastorAún no hay calificaciones
- Ecg CastellanosDocumento143 páginasEcg CastellanosVelia Gallegos100% (1)
- Biofisica CardíacaDocumento28 páginasBiofisica CardíacaMarcos Días100% (1)
- Marco Teorico ECGDocumento11 páginasMarco Teorico ECGEliana Caceres TorricoAún no hay calificaciones
- Anatomia y Fisiologia Del Sistema Excito ConductorDocumento16 páginasAnatomia y Fisiologia Del Sistema Excito Conductorrosica1973Aún no hay calificaciones
- Mod7 TaquiarritmiasDocumento73 páginasMod7 TaquiarritmiasFernando ArancibiaAún no hay calificaciones
- ElectrofisiologíaDocumento8 páginasElectrofisiologíaAndrea Valeria Montenegro Reyes100% (1)
- CARDIO - Bases Biofísicas de La Actividad Cardiaca y Músculo Liso Vascular Con Proyección A Bases FisiopatológicasDocumento28 páginasCARDIO - Bases Biofísicas de La Actividad Cardiaca y Músculo Liso Vascular Con Proyección A Bases FisiopatológicasRomina NogueiraAún no hay calificaciones
- Génesis Del ECG NormalDocumento8 páginasGénesis Del ECG NormalCamila CarpioAún no hay calificaciones
- Electrofisiologia PDFDocumento90 páginasElectrofisiologia PDFFabiola NorabuenaAún no hay calificaciones
- Practica Potencial de Acción CardiacoDocumento9 páginasPractica Potencial de Acción CardiacoXavi MGAún no hay calificaciones
- Origen y Propagación 2Documento9 páginasOrigen y Propagación 2Gabriela GregórioAún no hay calificaciones
- ARRITMIASDocumento21 páginasARRITMIASJoselyn RodriguezAún no hay calificaciones
- Seminario Tp4 Fisica BiologicaDocumento8 páginasSeminario Tp4 Fisica BiologicaEstefanía MansillaAún no hay calificaciones
- Potencial de Membrana y CardiovasculaDocumento7 páginasPotencial de Membrana y CardiovasculaJacki ServianAún no hay calificaciones
- 07 - Propiedades Del Músculo CardiacoDocumento11 páginas07 - Propiedades Del Músculo CardiacoAlejandra HernandezAún no hay calificaciones
- Clase 3 4-10Documento4 páginasClase 3 4-10Valentina UbillaAún no hay calificaciones
- CARDIACODocumento67 páginasCARDIACOMayreamar EstradaAún no hay calificaciones
- Electrocardiograma Normal y Patológico - Medicina InternaDocumento35 páginasElectrocardiograma Normal y Patológico - Medicina InternaPedro HermosoAún no hay calificaciones
- UntitledDocumento7 páginasUntitledZahira AbrahamAún no hay calificaciones
- Desfibrilador CardiacoDocumento46 páginasDesfibrilador CardiacoJesus Resta NavarroAún no hay calificaciones
- Ut II Guia10Documento9 páginasUt II Guia10BarbaraAndradeQuirozAún no hay calificaciones
- BiopotencialesDocumento20 páginasBiopotencialesRoland CaixaAún no hay calificaciones
- Fisiología Cardiovascular Electrocardiografía N°4 PDFDocumento22 páginasFisiología Cardiovascular Electrocardiografía N°4 PDFKevin Argotti CazarAún no hay calificaciones
- Mecanismos Electrofisiológicos de La Elevación Del Segmento ST Durante El Infarto Agudo de Miocardio. Hipótesis ActualDocumento7 páginasMecanismos Electrofisiológicos de La Elevación Del Segmento ST Durante El Infarto Agudo de Miocardio. Hipótesis ActualElíseo E LQAún no hay calificaciones
- Tarea 2 - Miguel Angel Ahumada Segura!!!Documento16 páginasTarea 2 - Miguel Angel Ahumada Segura!!!MIGUEL ANGEL AHUMADAAún no hay calificaciones
- Manual Toma e Interpretacion de ElectrocardiogramaDocumento29 páginasManual Toma e Interpretacion de ElectrocardiogramaGuillermina SanchezAún no hay calificaciones
- Aparato Cardiovascular 1T1Documento41 páginasAparato Cardiovascular 1T1Marycielo LmAún no hay calificaciones
- Fisiología - Guía # 1. Actividad Electrica Cardiaca 2023-2Documento10 páginasFisiología - Guía # 1. Actividad Electrica Cardiaca 2023-2LAURA SOFIA PEREZ CUELLOAún no hay calificaciones
- CARDIO!Documento31 páginasCARDIO!Mauricio VianaAún no hay calificaciones
- Propiedades Del Corazon de Sapo - CompressDocumento23 páginasPropiedades Del Corazon de Sapo - CompressJUAN RELUZ ZAMBRANOAún no hay calificaciones
- Electrofisiología CardíacaDocumento13 páginasElectrofisiología CardíacajohanaAún no hay calificaciones
- 1 EkgDocumento12 páginas1 EkgegleeAún no hay calificaciones
- Teoria Delm Musculo CardiacoDocumento12 páginasTeoria Delm Musculo CardiacoAlbert KantAún no hay calificaciones
- EKG Normal 1 y 2Documento12 páginasEKG Normal 1 y 2Franck FarreraAún no hay calificaciones
- GUÍA DE PRACTICA 3 Efecto de Los Iones Sobre La Frecuencia CardíacaDocumento4 páginasGUÍA DE PRACTICA 3 Efecto de Los Iones Sobre La Frecuencia CardíacaArely Mical SánchezAún no hay calificaciones
- Documento Sin TítuloDocumento15 páginasDocumento Sin TítuloGuillermina SanchezAún no hay calificaciones
- MielinizacionDocumento57 páginasMielinizacionVictoria Castillo100% (1)
- Electrofisiologia CardiacaDocumento14 páginasElectrofisiologia Cardiacaluispin1186Aún no hay calificaciones
- Electrofisiología y EcgDocumento11 páginasElectrofisiología y Ecgivanlalvarez.22Aún no hay calificaciones
- Automatismo AnormalDocumento3 páginasAutomatismo AnormalEmily Janice Oliva AlburquequeAún no hay calificaciones
- Propiedades Electricas Del CorazonDocumento4 páginasPropiedades Electricas Del CorazonRichard Armijo GarridoAún no hay calificaciones
- Secme-4237 1Documento11 páginasSecme-4237 1JoséAún no hay calificaciones
- Electrofisiologia Cardiaca CMDM PDFDocumento10 páginasElectrofisiologia Cardiaca CMDM PDFWendy OrtizAún no hay calificaciones
- Biofísica Cardiaca - YGDocumento3 páginasBiofísica Cardiaca - YGmichaellaAún no hay calificaciones
- BiopotencialesDocumento24 páginasBiopotencialesMiguel SoberanesAún no hay calificaciones
- Electrocardiograma IDocumento19 páginasElectrocardiograma IGeraldine LópezAún no hay calificaciones
- Tarea 2 - Trabajo ColaborativoDocumento46 páginasTarea 2 - Trabajo ColaborativoJuan Carlos Grisales MenesesAún no hay calificaciones
- Electro Fisio Log Í ADocumento33 páginasElectro Fisio Log Í Ajuancir2005jAún no hay calificaciones
- UTII 1 Practica Potencial de Accion CardiacoDocumento13 páginasUTII 1 Practica Potencial de Accion CardiacoMarisol RodríguezAún no hay calificaciones
- Cardiología CTODocumento95 páginasCardiología CTOEfren Penaflores100% (1)
- Apunte EelctrocardiografiaDocumento84 páginasApunte EelctrocardiografiaMelissa JerezAún no hay calificaciones
- Teoría-Función Cardiovascular - MEHU 632Documento24 páginasTeoría-Función Cardiovascular - MEHU 632Sara ParedesAún no hay calificaciones
- Electrocardiograma 01Documento15 páginasElectrocardiograma 01Yasirys Mariell PichardoAún no hay calificaciones
- (Estefany) Propiedades Electrofisiológicas Del Miocardio Contráctil y EspecializadoDocumento19 páginas(Estefany) Propiedades Electrofisiológicas Del Miocardio Contráctil y EspecializadoPIKAAún no hay calificaciones
- Cómo se comunican las neuronas: El milagro de la transmisión sinápticaDe EverandCómo se comunican las neuronas: El milagro de la transmisión sinápticaAún no hay calificaciones
- Medicina energética: Acupuntura 3 Circuitos energéticos secundariosDe EverandMedicina energética: Acupuntura 3 Circuitos energéticos secundariosCalificación: 5 de 5 estrellas5/5 (5)
- Informe 11.2 Codigo 530 UnaDocumento14 páginasInforme 11.2 Codigo 530 UnaAlicia SalinasAún no hay calificaciones
- Impresora 3D Prusa Mendel Iteration 2, Imprimiendo IdeasDocumento45 páginasImpresora 3D Prusa Mendel Iteration 2, Imprimiendo IdeasRomeo MuñozAún no hay calificaciones
- Semana 5 - Bienestar FetalDocumento5 páginasSemana 5 - Bienestar Fetaledith acosta castanedaAún no hay calificaciones
- Guia de Inmunología 2Documento5 páginasGuia de Inmunología 2Gabriela rojasAún no hay calificaciones
- Rebelion ZapatistaDocumento33 páginasRebelion ZapatistalavadodecerebroAún no hay calificaciones
- Métodos NuméricosDocumento16 páginasMétodos NuméricosfredyAún no hay calificaciones
- Reglamento Academico Aprobado El 24 de Mayo de 2018Documento24 páginasReglamento Academico Aprobado El 24 de Mayo de 2018MaternoAún no hay calificaciones
- Presas de Boveda MultipleDocumento13 páginasPresas de Boveda Multiplenehemias gomeroAún no hay calificaciones
- Preguntas NutricionDocumento3 páginasPreguntas NutricionRosii Gutiérrez VegaAún no hay calificaciones
- Uniendose A Shiva 1Documento142 páginasUniendose A Shiva 1Vanadis AlcyoneAún no hay calificaciones
- Farmacologia de Incontinencia Urinaria en El Adulto MayorDocumento26 páginasFarmacologia de Incontinencia Urinaria en El Adulto MayorJacqueline Vanessa Miranda SanabriaAún no hay calificaciones
- Boletín Censal 26Documento8 páginasBoletín Censal 26rosaleninAún no hay calificaciones
- Abuso Sexual. Documento de Vicaría EducaciónDocumento61 páginasAbuso Sexual. Documento de Vicaría EducaciónBrenda RossanaAún no hay calificaciones
- Resumen de Selección de PersonalDocumento17 páginasResumen de Selección de PersonalVictor Ariel GarayaldeAún no hay calificaciones
- Alergias RespiratoriasDocumento8 páginasAlergias RespiratoriasGabby Funes de Schaw100% (1)
- OVISEEEEEDocumento22 páginasOVISEEEEEJazmin CornejoAún no hay calificaciones
- Revista Rayo Verde 7Documento68 páginasRevista Rayo Verde 7Simon FalconiAún no hay calificaciones
- Derecho Laboral BTP BF 11moDocumento6 páginasDerecho Laboral BTP BF 11moEduardo SalandíaAún no hay calificaciones
- El ServidorDocumento12 páginasEl ServidorMariela SilgueroAún no hay calificaciones
- Caja Negra Caja Transparente Proceso deDocumento39 páginasCaja Negra Caja Transparente Proceso deLucero Alarcón100% (1)
- TENSOACTIVOSDocumento7 páginasTENSOACTIVOSAnonymous 3xmwtAaiuAún no hay calificaciones
- Informe de Laboratorio de Física ChoquesDocumento21 páginasInforme de Laboratorio de Física ChoquesCristianChancusiAún no hay calificaciones
- Evaluación 6°Documento30 páginasEvaluación 6°J Mario FQAún no hay calificaciones
- Metodo de Vicente ConesaDocumento14 páginasMetodo de Vicente ConesaGary Placencia100% (1)
- 1.2manual Exe LearningDocumento27 páginas1.2manual Exe LearningWilmer Tejada CabanillasAún no hay calificaciones
- Presentacion 1Documento15 páginasPresentacion 1Estrella Espinoza PaolaAún no hay calificaciones
- Cuadernillo Ciencias1.Biologia Sem2 (21-25 Sep)Documento17 páginasCuadernillo Ciencias1.Biologia Sem2 (21-25 Sep)Neto Kldra100% (1)
- Manual Kriyas y Med Primero 2016Documento188 páginasManual Kriyas y Med Primero 2016Carolina Ramírez100% (8)