También podría gustarte
- Resumen Fisiologia de Guyton Capitulo 40 Ed 12Documento6 páginasResumen Fisiologia de Guyton Capitulo 40 Ed 12XDXDXDXDAún no hay calificaciones
- NEUMODocumento35 páginasNEUMOLUZ ASTRID ELIGIO ANAYAAún no hay calificaciones
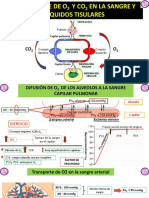
- 40 Transporte Del O2 y Co2 en La Sangre y Los Liquidos Tisulares Guyton 2 8 PDFDocumento7 páginas40 Transporte Del O2 y Co2 en La Sangre y Los Liquidos Tisulares Guyton 2 8 PDFAna RodriguezAún no hay calificaciones
- 004 Transporte de Oxigeno y Dioxido de CarbonoDocumento32 páginas004 Transporte de Oxigeno y Dioxido de CarbonoThulio MoutinhoAún no hay calificaciones
- Capitilo40 Transportedeo2yco2 121128200855 Phpapp02Documento42 páginasCapitilo40 Transportedeo2yco2 121128200855 Phpapp02Ginna Paola SaavedraAún no hay calificaciones
- Transporte de Oxígeno y Dióxido de Carbono en La Sangre y Líquidos TisularesDocumento5 páginasTransporte de Oxígeno y Dióxido de Carbono en La Sangre y Líquidos TisularesDayanara RodriguezAún no hay calificaciones
- Transporte e Intercambio Gaseoso. TextoDocumento7 páginasTransporte e Intercambio Gaseoso. TextoCarolina Rivera ReyAún no hay calificaciones
- Transporte de O2 y CO2 en La Sangre y Los Líquidos WORDDocumento6 páginasTransporte de O2 y CO2 en La Sangre y Los Líquidos WORDJessica Roman SejasAún no hay calificaciones
- Capitulo 41 Transporte de Oxigeno y Dioxido de Carbono en La Sangre y Los Liquidos Tisulares 96163 DownloableDocumento3 páginasCapitulo 41 Transporte de Oxigeno y Dioxido de Carbono en La Sangre y Los Liquidos Tisulares 96163 DownloableGiselle De La CruzAún no hay calificaciones
- Reporte de Lectura Capítulo 41 Transporte de Oxígeno y Dioxido de Carbono en La Sangre y Los Líquidos TisularesDocumento8 páginasReporte de Lectura Capítulo 41 Transporte de Oxígeno y Dioxido de Carbono en La Sangre y Los Líquidos TisularesMiguel CórdovaAún no hay calificaciones
- Resumen Transporte de GasesDocumento8 páginasResumen Transporte de GasesTefa 15Aún no hay calificaciones
- Transporte de Oxigeno de Los Pulmones A Los Tejidos Del OrganismoDocumento5 páginasTransporte de Oxigeno de Los Pulmones A Los Tejidos Del OrganismoEvelyn Calva0% (1)
- Difusión de Oxígeno de Los Capilares Periféricos Al Líquido TisularDocumento2 páginasDifusión de Oxígeno de Los Capilares Periféricos Al Líquido TisularStefany ChiricoAún no hay calificaciones
- PRÁCTICA 10 Transporte de Gases y Liquidos CorporalesDocumento13 páginasPRÁCTICA 10 Transporte de Gases y Liquidos CorporalesNoemi VelasquezAún no hay calificaciones
- CAPITULO-40 Fisiologia de Guyton - ResumenDocumento4 páginasCAPITULO-40 Fisiologia de Guyton - ResumenMiguel Angel Manotas CañasAún no hay calificaciones
- Capitulo 40Documento4 páginasCapitulo 40ArlenneRivera100% (2)
- Resumen Capitulo 41 Transporte O2 y Co2Documento5 páginasResumen Capitulo 41 Transporte O2 y Co2Lii LiiAún no hay calificaciones
- Cap 41 - Transporte de O2 y Co2 en La Sangre y Liquidos TisularesDocumento9 páginasCap 41 - Transporte de O2 y Co2 en La Sangre y Liquidos TisularesJennifer RiveraAún no hay calificaciones
- Capítulo 41 Guyton ResumenDocumento5 páginasCapítulo 41 Guyton ResumenJannely Nava MejíaAún no hay calificaciones
- Informe Transporte de O2 y CO2Documento5 páginasInforme Transporte de O2 y CO2Ruth Iraipi QuirogaAún no hay calificaciones
- Transporte de Oxígeno y Dióxido de Carbono en La Sangre y Líquidos Tisulares - Capítulo 41Documento7 páginasTransporte de Oxígeno y Dióxido de Carbono en La Sangre y Líquidos Tisulares - Capítulo 41Fernanda SotoAún no hay calificaciones
- Tema 4 Transporte de O2 y CO2 en La Sangre y Líquidos TisularesDocumento5 páginasTema 4 Transporte de O2 y CO2 en La Sangre y Líquidos TisularesAriana Gabriela Musaja Cruz100% (1)
- Transporte de OxigenoDocumento35 páginasTransporte de OxigenoGabuxita RodriguezAún no hay calificaciones
- Difusion de Gases Trans Barr Era HematogaseosaDocumento14 páginasDifusion de Gases Trans Barr Era HematogaseosaCarlos AfonsoAún no hay calificaciones
- Trabajo Extra - MergedDocumento26 páginasTrabajo Extra - MergedGeraldyne HernandezAún no hay calificaciones
- Transporte de O2 y Co2Documento8 páginasTransporte de O2 y Co2natAún no hay calificaciones
- Fisiologia - Respiratoria - III TRANSPORTE DE GASESDocumento29 páginasFisiologia - Respiratoria - III TRANSPORTE DE GASEScristian duqueAún no hay calificaciones
- Todo FisioDocumento30 páginasTodo FisioDoris RivadeneiraAún no hay calificaciones
- Transporte de O2 y CO2 en La Sangre y Líquidos TisularesDocumento43 páginasTransporte de O2 y CO2 en La Sangre y Líquidos Tisulareslucas herreraAún no hay calificaciones
- Transporte de Dioxido de Carbono y Oxigeno en La SangreDocumento30 páginasTransporte de Dioxido de Carbono y Oxigeno en La SangreEmilio Godoy MorenoAún no hay calificaciones
- Dr. Anibal Fisio II Capitulo 41Documento43 páginasDr. Anibal Fisio II Capitulo 41Bois de MariaAún no hay calificaciones
- Transporte Del Oxigeno y Dioxido de CarbonoDocumento29 páginasTransporte Del Oxigeno y Dioxido de CarbonoCristian CayoAún no hay calificaciones
- Ex PosicionDocumento2 páginasEx PosicionKarla Sánchez CórdovaAún no hay calificaciones
- Fisio Transporte de OxigenoDocumento18 páginasFisio Transporte de OxigenoAndrés MoraAún no hay calificaciones
- Introduccion Sistema Respiratorio PDFDocumento4 páginasIntroduccion Sistema Respiratorio PDFNicoleAún no hay calificaciones
- 29) Transporte de Gases en Sangre, Respiración CelularDocumento12 páginas29) Transporte de Gases en Sangre, Respiración Celularyulianaledesmantvg16Aún no hay calificaciones
- Tema 3 Introducción Fisiología PsicologíaDocumento5 páginasTema 3 Introducción Fisiología PsicologíaMartaAún no hay calificaciones
- Capitulo 41 - SebastiansanchezDocumento2 páginasCapitulo 41 - SebastiansanchezSebastian Sanchez MayaAún no hay calificaciones
- Perfusión 2021Documento17 páginasPerfusión 2021bdelkineAún no hay calificaciones
- Capitulo 41 Guyton H 393255 Downloadable 1374696Documento7 páginasCapitulo 41 Guyton H 393255 Downloadable 1374696ROY OMAR TINEO GUERRAAún no hay calificaciones
- Transporte de O2 y Co2Documento17 páginasTransporte de O2 y Co2carmen rosaAún no hay calificaciones
- Intercambio de Oxígeno y de Dióxido de CarbonoDocumento5 páginasIntercambio de Oxígeno y de Dióxido de CarbonoGalo Pinos100% (1)
- Act, 12 - Guadalupe Panama MenesesDocumento13 páginasAct, 12 - Guadalupe Panama MenesesGuadalupe PanamaAún no hay calificaciones
- F. Transporte de O2 y Co2 en La Sangre y Los Líquidos TisularesDocumento60 páginasF. Transporte de O2 y Co2 en La Sangre y Los Líquidos TisularesAlessia AndiaAún no hay calificaciones
- Intercambio de OxigenoDocumento22 páginasIntercambio de OxigenoSalvador BARRON MELOAún no hay calificaciones
- Sistema RespiratorioDocumento35 páginasSistema RespiratorioGABRIELA ALEJANDRA DE SANTIAGO LOPEZAún no hay calificaciones
- Difusión y Transporte de Gases en El EjercicioDocumento5 páginasDifusión y Transporte de Gases en El EjercicioDesiree LópezAún no hay calificaciones
- Cuidados de Enfermería A Usuarios Con Patologías RespiratoriasDocumento139 páginasCuidados de Enfermería A Usuarios Con Patologías RespiratoriasPablo Oddone100% (1)
- Sistema RespiratorioDocumento32 páginasSistema RespiratorioDaniel Montiel.Aún no hay calificaciones
- Capítulo 41Documento2 páginasCapítulo 41Juan Barcenas100% (1)
- Equilibrio Acido - BaseDocumento9 páginasEquilibrio Acido - BaseYuli HernandezAún no hay calificaciones
- Protocolo Ventilacion Pulmonar 1Documento20 páginasProtocolo Ventilacion Pulmonar 1Ruth Esther Nuñez MaldonadoAún no hay calificaciones
- Biofisica de La RespiraciónDocumento19 páginasBiofisica de La RespiraciónemmanuelAún no hay calificaciones
- Tema 26Documento4 páginasTema 26gabrielmd858Aún no hay calificaciones
- Cianosis PDFDocumento6 páginasCianosis PDFGissSánchezAún no hay calificaciones
- Clase IxDocumento22 páginasClase IxesthersitapacoAún no hay calificaciones
- Relacion Ventilación - PerfusiónDocumento11 páginasRelacion Ventilación - PerfusiónJosmirys Rendon SifontesAún no hay calificaciones
- Cuestionario Sistema RESPIRATORIODocumento3 páginasCuestionario Sistema RESPIRATORIOSARAY TREJO ROQUEAún no hay calificaciones
- Conceptos Y Estrategias En Terapia RespiratoriaDe EverandConceptos Y Estrategias En Terapia RespiratoriaCalificación: 4 de 5 estrellas4/5 (1)
- Farm Acolo GiaDocumento48 páginasFarm Acolo GiaCamila OrmeñoAún no hay calificaciones
- Introducción A Las Baterías de LitioDocumento27 páginasIntroducción A Las Baterías de LitioSORAYA MARTINEZ MAYGUAAún no hay calificaciones
- PoliflexDocumento16 páginasPoliflexjose_arqtovAún no hay calificaciones
- Ejemplo AAADocumento7 páginasEjemplo AAAhalina pradaAún no hay calificaciones
- Informe de Celula VegetalDocumento7 páginasInforme de Celula VegetalÑzñz Redes75% (4)
- Proyecto de QuimicaDocumento3 páginasProyecto de QuimicaGael GarciaAún no hay calificaciones
- Ejer1 MNDocumento3 páginasEjer1 MNjjacccobinoAún no hay calificaciones
- Cultivo de AlgasDocumento34 páginasCultivo de AlgasBrenda Fabiola Chavesta100% (1)
- Guía de Físicoquimica - Sexto GradoDocumento2 páginasGuía de Físicoquimica - Sexto GradoJuan Martin Sierra CarrascalAún no hay calificaciones
- Transaminacion - Secc 3Documento2 páginasTransaminacion - Secc 3nicole jacomeAún no hay calificaciones
- Propiedades Químicas Del SueloDocumento52 páginasPropiedades Químicas Del SueloKarlo FernandoAún no hay calificaciones
- Lectura de HologramasDocumento12 páginasLectura de HologramasJohnny Jeremias Gutierrez Martinez0% (1)
- Síntesis de P2NP Catalizador de Acetato de AmonioDocumento2 páginasSíntesis de P2NP Catalizador de Acetato de AmonioMoises Valdivia BaldomeroAún no hay calificaciones
- Libro Practicas Procesos de Tejidos IiDocumento69 páginasLibro Practicas Procesos de Tejidos IiEncarnaAún no hay calificaciones
- Oxido de EstirenoDocumento6 páginasOxido de EstirenoMoises Valdivia BaldomeroAún no hay calificaciones
- Practica Diagnostico Del Estado Del Motor Por Medio Del Analisis de Los GasesDocumento7 páginasPractica Diagnostico Del Estado Del Motor Por Medio Del Analisis de Los Gaseswilliam_senati20057051Aún no hay calificaciones
- D6 CarmixDocumento2 páginasD6 CarmixEVCYAún no hay calificaciones
- 04 911509yzjagoncph PDFDocumento43 páginas04 911509yzjagoncph PDFReinaldo Quiroga MendozaAún no hay calificaciones
- Nombres Comunes de HeterociclosDocumento5 páginasNombres Comunes de HeterociclosMilagros RSAún no hay calificaciones
- Sistema de AlimentaciónDocumento25 páginasSistema de AlimentaciónRene MariscalAún no hay calificaciones
- Especificación Técnica #P - 152Documento9 páginasEspecificación Técnica #P - 152PROYECTOMSAún no hay calificaciones
- Práctica 06 - Método Del Número Más Problable Pra La Identificación de Microorganismos en Alimentos.Documento7 páginasPráctica 06 - Método Del Número Más Problable Pra La Identificación de Microorganismos en Alimentos.Stephanie ZapataAún no hay calificaciones
- TrituracionDocumento24 páginasTrituracionjasinto_222Aún no hay calificaciones
- Reporte 14 QgiiDocumento5 páginasReporte 14 QgiiGab OCAún no hay calificaciones
- Lab Siderurgia DavalosDocumento14 páginasLab Siderurgia Davalosrubenmanuel5Aún no hay calificaciones
- Guía de Estudio - Lodo Activo - DiseñoDocumento6 páginasGuía de Estudio - Lodo Activo - DiseñoCarol VidalAún no hay calificaciones
- Linus Carl PaulingDocumento16 páginasLinus Carl PaulingJose LugoAún no hay calificaciones
- Manual ViledonDocumento2 páginasManual ViledonDaniel RomeroAún no hay calificaciones
- Manual Fluidos de Perforación - 000Documento792 páginasManual Fluidos de Perforación - 000Julio RucheAún no hay calificaciones
- Anemia y Sangre de Pollo OpcionalDocumento5 páginasAnemia y Sangre de Pollo Opcionalmedical9Aún no hay calificaciones