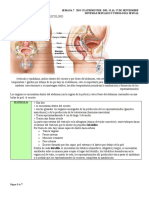
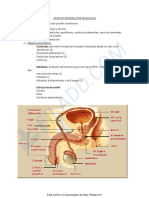
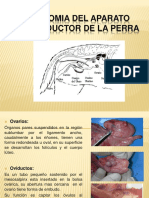
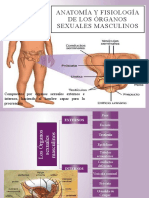
También podría gustarte
- TRIPTICO Aparato ReproductorDocumento2 páginasTRIPTICO Aparato ReproductorGloria Villacorta77% (13)
- Fisiologia Del Ciclo SexualDocumento24 páginasFisiologia Del Ciclo SexualArgelia OG100% (2)
- Aparato Genital MasculinoDocumento30 páginasAparato Genital MasculinoCristina Montalvo SolsAún no hay calificaciones
- Marco Teórico: Generalidades Del Aparato Reproductor MasculinoDocumento18 páginasMarco Teórico: Generalidades Del Aparato Reproductor MasculinoAndrea IriasAún no hay calificaciones
- 3er Parcial EndoDocumento7 páginas3er Parcial EndoCarolina Valdez RuizAún no hay calificaciones
- Sistema Reproductor Masculino6originalDocumento20 páginasSistema Reproductor Masculino6originalCristian CamposAún no hay calificaciones
- TESTICULO1Documento18 páginasTESTICULO1Natalia VergaraAún no hay calificaciones
- Clase Semana 13 - Aparato Reproductor MasculinoDocumento36 páginasClase Semana 13 - Aparato Reproductor MasculinoMarycielo LmAún no hay calificaciones
- Los TestículosDocumento11 páginasLos TestículosAndres BolivarAún no hay calificaciones
- Resumen de Aparato Reproductor Masculino DesiDocumento7 páginasResumen de Aparato Reproductor Masculino DesiDaya GarciaAún no hay calificaciones
- Sitema Reproductor MasculinoDocumento16 páginasSitema Reproductor MasculinoJesus Mollinedo mrenoAún no hay calificaciones
- Sistema Reproductor MasculinoDocumento6 páginasSistema Reproductor MasculinoJahaziel MejiaAún no hay calificaciones
- Aparato Reproductor MasculinoDocumento11 páginasAparato Reproductor MasculinoNair MartínezAún no hay calificaciones
- Sistema Reproductor MasculinoDocumento135 páginasSistema Reproductor MasculinoPaola Azucena SalasAún no hay calificaciones
- HISTOLOGIADocumento7 páginasHISTOLOGIAmellisa olivares villaAún no hay calificaciones
- AP - Reproductor MasculinoDocumento12 páginasAP - Reproductor MasculinoantonellamartAún no hay calificaciones
- FUNCIONES REPRODUCTORAS Y HORMONALES MASCULINAS ResumenDocumento9 páginasFUNCIONES REPRODUCTORAS Y HORMONALES MASCULINAS ResumenIvettPaolaSamaniegoAún no hay calificaciones
- Sistemas SexualesDocumento7 páginasSistemas SexualesAinara zubillagaAún no hay calificaciones
- Sistema Reproductor Masculino (1) ORIGINALDocumento24 páginasSistema Reproductor Masculino (1) ORIGINALCristian CamposAún no hay calificaciones
- Aparato Reproductor Masculino 3Documento17 páginasAparato Reproductor Masculino 3Juliana GonzálezAún no hay calificaciones
- Embio P3Documento14 páginasEmbio P3ERIKA VALERIA YAR NAULAAún no hay calificaciones
- Masculoino K PDFDocumento18 páginasMasculoino K PDFKimberly ReyesAún no hay calificaciones
- Brenda Lizet Hernández Sandoval Odontología 2B Patología General Mtra. María Dolores Cecilia Gonzales MalagónDocumento8 páginasBrenda Lizet Hernández Sandoval Odontología 2B Patología General Mtra. María Dolores Cecilia Gonzales MalagónBrenda Lizet Hernandez SandovalAún no hay calificaciones
- Tema 39 Reducido A 19 PaginasDocumento19 páginasTema 39 Reducido A 19 PaginasAngel CardenasAún no hay calificaciones
- Aparato Reproducto Masculino 23Documento30 páginasAparato Reproducto Masculino 23andrezmendez09Aún no hay calificaciones
- Aparato Reproductor MasculinoDocumento7 páginasAparato Reproductor MasculinoLucia Nieva AllueAún no hay calificaciones
- GAMETOGÉNESISDocumento7 páginasGAMETOGÉNESISMiguel Angel Parra GarcíaAún no hay calificaciones
- Sistema Reproductor - Ap Genital MasculinoDocumento5 páginasSistema Reproductor - Ap Genital Masculinorociofigueroa.m130Aún no hay calificaciones
- Hipogonadismo Masculino y SD de KlinefelterDocumento40 páginasHipogonadismo Masculino y SD de KlinefelterAnghi León HidalgoAún no hay calificaciones
- Anatomia y Fisiologia Sistema Reproductor Masculino Uap 2015Documento60 páginasAnatomia y Fisiologia Sistema Reproductor Masculino Uap 2015Joshelin GonzalesAún no hay calificaciones
- Testículos HISTOLOGIADocumento8 páginasTestículos HISTOLOGIACindy CuadrosAún no hay calificaciones
- Orientadoras 11 y 12 Morfofisiologia Humana 3Documento25 páginasOrientadoras 11 y 12 Morfofisiologia Humana 3Dayerlin HernandezAún no hay calificaciones
- Biologia Meosis y MitosisDocumento8 páginasBiologia Meosis y MitosisjimenaAún no hay calificaciones
- Embriogénesis Del Aparato Genital MasculinoDocumento17 páginasEmbriogénesis Del Aparato Genital MasculinoAna Cristina Sanchez ZangAún no hay calificaciones
- Aparato Reproductor MasculinoDocumento5 páginasAparato Reproductor MasculinoIzbeth TorresAún no hay calificaciones
- 2.2) Masculino PDFDocumento6 páginas2.2) Masculino PDFGiaan OrtizAún no hay calificaciones
- Aparato Reproductor FemeninoDocumento43 páginasAparato Reproductor FemeninoKarin Montenegro DiazAún no hay calificaciones
- Anatomia y Fisiologia Del Aparato Reproductor Masculino ResumenDocumento5 páginasAnatomia y Fisiologia Del Aparato Reproductor Masculino ResumenYessenia CatterineAún no hay calificaciones
- Aparatos ReproductoresDocumento38 páginasAparatos ReproductoresAlisson CahuanaAún no hay calificaciones
- Aparato Reproductor MasculinoDocumento4 páginasAparato Reproductor MasculinoArmando Martínez CanoAún no hay calificaciones
- ApuntesDocumento7 páginasApuntesJoseline Pineda GarcíaAún no hay calificaciones
- REPRODUCCIONDocumento31 páginasREPRODUCCIONDuvan SanabriaAún no hay calificaciones
- ReproductoresDocumento28 páginasReproductoresAylen KesslerAún no hay calificaciones
- Sistema Reproductor Masculino. InformeDocumento10 páginasSistema Reproductor Masculino. Informemanuel quintanillaAún no hay calificaciones
- E SpermatogenesisDocumento6 páginasE SpermatogenesisThalía YanchapaxiAún no hay calificaciones
- Reproductor MyFDocumento103 páginasReproductor MyFJosué Antonio Morales LorioAún no hay calificaciones
- Embriologia Sistema Genital PDFDocumento11 páginasEmbriologia Sistema Genital PDFJOCELYN FIERRO GUERREROAún no hay calificaciones
- Aparato Reproductor Masculino TejidosDocumento11 páginasAparato Reproductor Masculino Tejidospanther100Aún no hay calificaciones
- Aparato Reproductor Masculino:: ÓrganosDocumento6 páginasAparato Reproductor Masculino:: Órganoskim taehyungAún no hay calificaciones
- Histología Del Aparato Reproductor MasculinoDocumento14 páginasHistología Del Aparato Reproductor Masculinogonzalezruizandres500Aún no hay calificaciones
- TestículoDocumento36 páginasTestículoJesusAntonioAún no hay calificaciones
- FISIOLOGIA Aparato Reproductor MasculinoDocumento13 páginasFISIOLOGIA Aparato Reproductor MasculinoStefy AguaizaAún no hay calificaciones
- Histologia Del TesticuloDocumento20 páginasHistologia Del TesticuloHugo Richard Flores ContrerasAún no hay calificaciones
- ReproducciónDocumento10 páginasReproducciónDonatella AcuñaAún no hay calificaciones
- 1 BASES FISIOLOGICAS DE LA REPRODUCCION EN EL MACHO SeparataDocumento5 páginas1 BASES FISIOLOGICAS DE LA REPRODUCCION EN EL MACHO SeparataEstefania LN VasquezAún no hay calificaciones
- Taller de Reproducción HumanaDocumento24 páginasTaller de Reproducción HumanaMaryoris Del C. Osorio DE LA OssaAún no hay calificaciones
- Guia 1 Morfologia Del FdgreproductivoDocumento12 páginasGuia 1 Morfologia Del FdgreproductivoAlvaroJJaimesCAún no hay calificaciones
- Anatomía y Embriología de Los TesticulosDocumento18 páginasAnatomía y Embriología de Los TesticulosLoOgan2394100% (3)
- Clase Histologia MasculinaDocumento59 páginasClase Histologia MasculinaJorge Enrique Vera SagásteguiAún no hay calificaciones
- Reproducción Sexual en Humanos. Parte 1Documento6 páginasReproducción Sexual en Humanos. Parte 1pokeraperoAún no hay calificaciones
- 11 Aparato Reproductor MasculinoDocumento32 páginas11 Aparato Reproductor MasculinoYocelin GUTIERREZAún no hay calificaciones
- Anatomia y Fisiologia Del Aparato ReproductorrDocumento37 páginasAnatomia y Fisiologia Del Aparato ReproductorrDaniel Dreyfus AnguloAún no hay calificaciones
- Evaluacion Caracteristicas Microscopicas Del Semen PorcinoDocumento13 páginasEvaluacion Caracteristicas Microscopicas Del Semen PorcinoDiego DelgadoAún no hay calificaciones
- EXAMEN 1er BIMESTRE Biologia SecundariaDocumento5 páginasEXAMEN 1er BIMESTRE Biologia SecundariaBeto Junagut GavgAún no hay calificaciones
- Anatomía y Fisiología de Los Órganos Sexuales MasculinosDocumento10 páginasAnatomía y Fisiología de Los Órganos Sexuales MasculinosjosephAún no hay calificaciones
- CRIPTORQUIDIADocumento50 páginasCRIPTORQUIDIANorita Ruiz PalaciosAún no hay calificaciones
- Anatomia Genitales Externos FemeninosDocumento10 páginasAnatomia Genitales Externos Femeninosdesiree alvarado torgeAún no hay calificaciones
- Metabolismo de Una FármacoDocumento7 páginasMetabolismo de Una FármacoMiranda SPAún no hay calificaciones
- Aparato Reproductor MasculinoDocumento23 páginasAparato Reproductor MasculinoBeatriz Milagros PeveAún no hay calificaciones
- Anatomia Uretra Masculina y FemeninaDocumento21 páginasAnatomia Uretra Masculina y FemeninaCarlos Alberto Solorzano Suriaga100% (2)
- Ciclo OvaricoDocumento13 páginasCiclo OvaricoEscarla FructuosoAún no hay calificaciones
- Trabajo de ReproduccionDocumento26 páginasTrabajo de ReproduccionKaren OrtizAún no hay calificaciones
- Anatomia y FisiologiaDocumento14 páginasAnatomia y FisiologiaGarcía JosuéAún no hay calificaciones
- CAPÍTULO 20 - Sistema Reproductor Masculino-1Documento15 páginasCAPÍTULO 20 - Sistema Reproductor Masculino-1ANDREA ALEJANDRA OCHOA NAVARROAún no hay calificaciones
- Varicoccele (Autoguardado)Documento28 páginasVaricoccele (Autoguardado)ZENTENOAún no hay calificaciones
- Producto de CienciasDocumento10 páginasProducto de CienciasAdriano De Los Santos FloresAún no hay calificaciones
- Palpacion Genital MasculinoDocumento1 páginaPalpacion Genital MasculinoMedicina UnisimonAún no hay calificaciones
- Aparato Reproductor Femenino - 5Documento6 páginasAparato Reproductor Femenino - 5monique violliAún no hay calificaciones
- Tríptico Edu. Sex.Documento2 páginasTríptico Edu. Sex.IoselyneAún no hay calificaciones
- FimosisDocumento8 páginasFimosislucia mayaAún no hay calificaciones
- Triptico Aparato Reproductor MasculinoDocumento2 páginasTriptico Aparato Reproductor MasculinoMARIA DOMINGUEZ RIVERA0% (1)
- PPPPPPPPPPPPPPPPPPPPPPPDocumento40 páginasPPPPPPPPPPPPPPPPPPPPPPPdulceAún no hay calificaciones
- Semiología Aparato Reproductor FemeninoDocumento27 páginasSemiología Aparato Reproductor FemeninoJose Juan GalavizAún no hay calificaciones
- Sem 14 Embriologia Ap. Femenino 2021 1Documento46 páginasSem 14 Embriologia Ap. Femenino 2021 1Yahaira NeyraAún no hay calificaciones
- Universidad Católica de Santiago de Guayaquil Facultad de Ciencias Médicas. Carrera de MedicinaDocumento4 páginasUniversidad Católica de Santiago de Guayaquil Facultad de Ciencias Médicas. Carrera de Medicinakeyla Lastra BermeoAún no hay calificaciones
- Sistema Reproductor FemeninoDocumento56 páginasSistema Reproductor FemeninoKevin Flores100% (4)
- Aparato Genital MasculinoDocumento6 páginasAparato Genital MasculinoDiego Raul Davila VargasAún no hay calificaciones
- GenitalesDocumento26 páginasGenitalessinellyAún no hay calificaciones
- Ciclo Menstrual, Ovarico, EndometrialDocumento5 páginasCiclo Menstrual, Ovarico, EndometrialAndrés CervantesAún no hay calificaciones