También podría gustarte
- MF0712_1 - Operaciones auxiliares en reproducción ganaderaDe EverandMF0712_1 - Operaciones auxiliares en reproducción ganaderaAún no hay calificaciones
- Efecto MachoDocumento9 páginasEfecto MachoAlirio Villanova GonzalezAún no hay calificaciones
- Actividad 2 - EmbriologíaDocumento5 páginasActividad 2 - EmbriologíaLAURA MARIA BUSTOS VILLEGASAún no hay calificaciones
- Aparato Reproductor Femenino ConejosDocumento5 páginasAparato Reproductor Femenino ConejosRosa Amelia Marcelo MairaAún no hay calificaciones
- III - Unidad - Fecundacion - Segmentacion 2020Documento4 páginasIII - Unidad - Fecundacion - Segmentacion 2020Michelle CatalánAún no hay calificaciones
- TALLERDocumento11 páginasTALLERandres contreras ramirezAún no hay calificaciones
- 12 Sistema Reproductor HembraDocumento24 páginas12 Sistema Reproductor HembraMarco Antonio PeñaAún no hay calificaciones
- Fisiología Sexual en CaninosDocumento7 páginasFisiología Sexual en CaninosmacroposeAún no hay calificaciones
- Inseminacion Artificial en PerrasDocumento15 páginasInseminacion Artificial en PerrasMaría Paz Ramos YateAún no hay calificaciones
- Reproduccion HumanaDocumento6 páginasReproduccion HumanaMichell ReyesAún no hay calificaciones
- Tipeos Repro 2Documento60 páginasTipeos Repro 2Pía TroncosoAún no hay calificaciones
- Exposicion de Reproduccion de PerrosDocumento18 páginasExposicion de Reproduccion de PerrosCarmelita Hcs RmrezAún no hay calificaciones
- 03-Ia CabrasDocumento17 páginas03-Ia CabrasMary MartinezAún no hay calificaciones
- PIA (Producto Integrador de Aprendizaje) Reproducción Animal "Ciclo Estral de La Perra"Documento20 páginasPIA (Producto Integrador de Aprendizaje) Reproducción Animal "Ciclo Estral de La Perra"Dani RosalesAún no hay calificaciones
- Informe de Citologia Vaginal en PerrasDocumento11 páginasInforme de Citologia Vaginal en PerrasKaren Arancibia Fernández100% (1)
- Pubertad II UnidadDocumento17 páginasPubertad II UnidadGuillermo AguilarAún no hay calificaciones
- Transporte EspermáticoDocumento6 páginasTransporte EspermáticoTatiana RoldánAún no hay calificaciones
- Reproduccion Bovina SenaDocumento29 páginasReproduccion Bovina SenaPedro José RojasAún no hay calificaciones
- Ciclo Estral de La CabraDocumento7 páginasCiclo Estral de La Cabraleonel milanoAún no hay calificaciones
- Guia de Reproduccion y GestacionDocumento10 páginasGuia de Reproduccion y GestacionJose Alejandro Velasco MondragonAún no hay calificaciones
- Anatomía y Fisiología Del Tracto Reproductivo de La Hembra y Del Macho BovinoDocumento5 páginasAnatomía y Fisiología Del Tracto Reproductivo de La Hembra y Del Macho BovinoAlirio Villanova Gonzalez0% (1)
- Parametros y Productivos de Las AvesDocumento12 páginasParametros y Productivos de Las AvesEliseo Carpio50% (2)
- Fisiologia TurboDocumento3 páginasFisiologia Turbosilvio latiniAún no hay calificaciones
- INFORME 02 - Ciclo Estral y Control Hormonal en Las Especies Animales (Perra, Vaca, Yegua, Cabra y Cuy) .Documento25 páginasINFORME 02 - Ciclo Estral y Control Hormonal en Las Especies Animales (Perra, Vaca, Yegua, Cabra y Cuy) .NATALI MANUELA ZEGARRA REA??OAún no hay calificaciones
- Fisiologia de La Reproduccion en Aves Domesticas PDFDocumento26 páginasFisiologia de La Reproduccion en Aves Domesticas PDFCarlos Saucedo75% (4)
- Aparato Reproductor de La Hembra BovinaDocumento17 páginasAparato Reproductor de La Hembra BovinaYesely bastidas100% (1)
- Fisiología de La Reproducción de Los Animales DomésticosDocumento8 páginasFisiología de La Reproducción de Los Animales DomésticosElvis DugarteAún no hay calificaciones
- Repro Parcial IIDocumento47 páginasRepro Parcial IILisneidy RoaAún no hay calificaciones
- Ciclo Estral y Analisis SeminalDocumento9 páginasCiclo Estral y Analisis SeminalMisael Jake OrtizAún no hay calificaciones
- Mula RosaritoDocumento13 páginasMula RosaritoIsaacBarreraAún no hay calificaciones
- Reproduccion de Ovinos y CaprinosDocumento16 páginasReproduccion de Ovinos y CaprinosErick CanelónAún no hay calificaciones
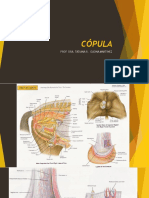
- Cópula PDFDocumento49 páginasCópula PDFAracely SanchezAún no hay calificaciones
- Ciclo AstralDocumento6 páginasCiclo AstralJesus RamirezAún no hay calificaciones
- Fisisologia Reproductiva Del ConejoDocumento8 páginasFisisologia Reproductiva Del ConejoDANIELA UNTUÑAAún no hay calificaciones
- Fisiologia de La ReproduccionDocumento20 páginasFisiologia de La Reproduccionadobecreativecloudcp1Aún no hay calificaciones
- Reproduccion en Ovejas ?Documento9 páginasReproduccion en Ovejas ?Luis angel LlancoAún no hay calificaciones
- Fecundacion Hmi FinalDocumento21 páginasFecundacion Hmi FinalDaniela Gutiérrez AlanísAún no hay calificaciones
- Reproduccion en OvinosDocumento38 páginasReproduccion en OvinosZavala H IrvingAún no hay calificaciones
- Ciclo Estral de La PerraDocumento29 páginasCiclo Estral de La PerraPamela Alvarez de MendozaAún no hay calificaciones
- Caninos y FelinosDocumento19 páginasCaninos y FelinosFernanda Miranda RiberaAún no hay calificaciones
- Manual de Reproducción BovinaDocumento20 páginasManual de Reproducción BovinaGabriel MoyaAún no hay calificaciones
- Citologia VaginalDocumento20 páginasCitologia VaginalArnold David Requena MartensAún no hay calificaciones
- Ciclos Reproductores y Continuos PDFDocumento3 páginasCiclos Reproductores y Continuos PDFKlaudiia VenegasAún no hay calificaciones
- Desarrollo EmbrionarioDocumento16 páginasDesarrollo EmbrionarioAllenWalkerAún no hay calificaciones
- Transporte y Sobrevivencia de Los GametosDocumento22 páginasTransporte y Sobrevivencia de Los GametosjasminAún no hay calificaciones
- Ciclo Estral de Los AnimalesDocumento7 páginasCiclo Estral de Los AnimalesSabino Vega GonzalesAún no hay calificaciones
- Sistema Reproductor - Ap Genital FemeninoDocumento10 páginasSistema Reproductor - Ap Genital Femeninorociofigueroa.m130Aún no hay calificaciones
- Cortejo y Apareamiento en 4 Especies de Animales Domésticos.Documento7 páginasCortejo y Apareamiento en 4 Especies de Animales Domésticos.Jaqueline Guerrero FloresAún no hay calificaciones
- Reproduccion BovinaDocumento4 páginasReproduccion BovinaRuben AcevedoAún no hay calificaciones
- Anatomía y Fisiología Del Tracto Reproductivo de La Hembra y Del Macho BovinoDocumento28 páginasAnatomía y Fisiología Del Tracto Reproductivo de La Hembra y Del Macho BovinoSAMUEL ALFONSO GUZMÁN DÍAZAún no hay calificaciones
- Taller de Bovinos Reproduccion de La Hembra BovinaDocumento8 páginasTaller de Bovinos Reproduccion de La Hembra BovinaEstudiante VeterinarioAún no hay calificaciones
- Repro Caninos Parte 1 PDFDocumento27 páginasRepro Caninos Parte 1 PDFantonioAún no hay calificaciones
- Fisiologia Del GalloDocumento9 páginasFisiologia Del GalloEliza LiraAún no hay calificaciones
- Mecanismo de RegulacionDocumento29 páginasMecanismo de Regulacionyacson cano valenciaAún no hay calificaciones
- Semiologia Aparato Reproductor HembraDocumento20 páginasSemiologia Aparato Reproductor Hembradanze33% (3)
- Formación de Los Folículos en El OvarioDocumento7 páginasFormación de Los Folículos en El OvarioAdriian NaviaAún no hay calificaciones
- Fisiologia ReproductivaDocumento44 páginasFisiologia ReproductivaCesar Max Perlacios VargasAún no hay calificaciones
- Reproduccion BovinaDocumento21 páginasReproduccion BovinaIselen MejiaAún no hay calificaciones
- Órgano VestibulococlearDocumento10 páginasÓrgano VestibulococlearLucaAún no hay calificaciones
- Fisiología de Rumiantes (Final)Documento17 páginasFisiología de Rumiantes (Final)LucaAún no hay calificaciones
- MeningesDocumento2 páginasMeningesLucaAún no hay calificaciones
- Estructura BacterianaDocumento9 páginasEstructura BacterianaLucaAún no hay calificaciones
- Órgano de La VisiónDocumento14 páginasÓrgano de La VisiónLucaAún no hay calificaciones
- Cavidad AbdominalDocumento1 páginaCavidad AbdominalLucaAún no hay calificaciones
- Inmunidad Activa Artificial UNLPDocumento29 páginasInmunidad Activa Artificial UNLPLucaAún no hay calificaciones
- Ficha 8 MatemáticaDocumento12 páginasFicha 8 MatemáticaJuan Carlos MadaAún no hay calificaciones
- Criterio Financieros para Seleccion de InversionesDocumento49 páginasCriterio Financieros para Seleccion de InversionesAsdfAún no hay calificaciones
- Seguridad Activa y PasivaDocumento2 páginasSeguridad Activa y PasivaJhonnatan AldasAún no hay calificaciones
- Cuarzo Ceramico M200Documento1 páginaCuarzo Ceramico M200racupi3Aún no hay calificaciones
- Impacto Del FEFODocumento6 páginasImpacto Del FEFOPatricia Condezo MárquezAún no hay calificaciones
- Retie 2010Documento22 páginasRetie 2010Alexander FraileAún no hay calificaciones
- INSTRUMENTOSDocumento13 páginasINSTRUMENTOSJorge Victor Urbina CruzAún no hay calificaciones
- Et200sp Io Link Master Manual es-ES es-ESDocumento42 páginasEt200sp Io Link Master Manual es-ES es-ESKelly Perez LopezAún no hay calificaciones
- TEMA 5 Arte Paleocristiano y Bizantino PDFDocumento14 páginasTEMA 5 Arte Paleocristiano y Bizantino PDFzarandaja1100% (1)
- 3.2 PSFDocumento43 páginas3.2 PSFQuirino Suri100% (1)
- Clave Tipos de LarvaDocumento1 páginaClave Tipos de LarvaDaniel Felipe Morera LopezAún no hay calificaciones
- Las Tentaciones de Los Lideres - P Benigno JuanesDocumento43 páginasLas Tentaciones de Los Lideres - P Benigno JuanesEduardo Baca Contreras100% (7)
- PFC Mario Simon BordonDocumento102 páginasPFC Mario Simon BordonSteven Michael Rodríguez CaveroAún no hay calificaciones
- Programa Nacional Integral de Educación y Seguridad VialDocumento3 páginasPrograma Nacional Integral de Educación y Seguridad VialEleazarClavoAún no hay calificaciones
- GitlabDocumento5 páginasGitlabVanessa MendozaAún no hay calificaciones
- Micosis Sist HospDocumento78 páginasMicosis Sist HospDayanaAún no hay calificaciones
- C V PJHMDocumento4 páginasC V PJHMJose MamaniAún no hay calificaciones
- Formato PlaceDocumento11 páginasFormato PlaceKarenAún no hay calificaciones
- Tema Tesis UdchDocumento16 páginasTema Tesis UdchJoseSilvaAún no hay calificaciones
- Tabla de Convalidacion - Ing - Ind - Pensum 2016Documento1 páginaTabla de Convalidacion - Ing - Ind - Pensum 2016Sobeida JimenezAún no hay calificaciones
- Situacion Actual de La Produccion de Cambur en El Estado AraguaDocumento42 páginasSituacion Actual de La Produccion de Cambur en El Estado AraguaTomasVAún no hay calificaciones
- Ensayo de Carga Con PlacaDocumento10 páginasEnsayo de Carga Con PlacaBrishely Luis CondeAún no hay calificaciones
- CuarzoDocumento21 páginasCuarzoJesus ChaucaAún no hay calificaciones
- HM - Maratón de Las Fijas Acem 4-10-2022Documento8 páginasHM - Maratón de Las Fijas Acem 4-10-2022Laura Diana Quispe IngaAún no hay calificaciones
- Almacenamiento de Granos Verdes - Factores Que Debes Controlar - Perfect Daily GrindDocumento25 páginasAlmacenamiento de Granos Verdes - Factores Que Debes Controlar - Perfect Daily GrindLuis Antonio RiosAún no hay calificaciones
- 8 Triptico Uso de ExtintoresDocumento2 páginas8 Triptico Uso de ExtintoresMarAg KaragozAún no hay calificaciones
- Tesis Tajo Raul RojasDocumento154 páginasTesis Tajo Raul RojasTHALIA YAJAIRA RIMAC VEGAAún no hay calificaciones
- Residuos Sólidos y Consumo Sustentable PDFDocumento100 páginasResiduos Sólidos y Consumo Sustentable PDFYoselin CalderónAún no hay calificaciones
- EXPOSICION Auditoria 2019Documento11 páginasEXPOSICION Auditoria 2019JhonerValdezValdivianoAún no hay calificaciones
- Clase 2.2 Clasificación de Las Rocas 25 05 22Documento37 páginasClase 2.2 Clasificación de Las Rocas 25 05 22Pame JácomeAún no hay calificaciones