También podría gustarte
- Biotecnología y Alimentos FuncionalesDocumento28 páginasBiotecnología y Alimentos FuncionalesIsrael BimbozaAún no hay calificaciones
- Nutrient EsDocumento7 páginasNutrient EsLuzhesita Marix Martinez RiosAún no hay calificaciones
- CARBOHIDRATOSDocumento16 páginasCARBOHIDRATOSkeisy AracheAún no hay calificaciones
- Diabetes Gestacional Smne PDFDocumento2 páginasDiabetes Gestacional Smne PDFLuciaGomezAún no hay calificaciones
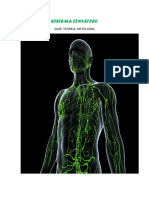
- Sistema LinfaticoDocumento14 páginasSistema LinfaticokellyvanessamendozahuamanAún no hay calificaciones
- Universidad Nacional Pedro Ruiz GalloDocumento9 páginasUniversidad Nacional Pedro Ruiz GalloAngela TocasSilvaAún no hay calificaciones
- Sistema Celular InteiDocumento55 páginasSistema Celular InteiIsabela MartinezAún no hay calificaciones
- Libro CardiovascularDocumento103 páginasLibro CardiovascularElizabeth AtencioAún no hay calificaciones
- Informe Paradigma Cualitativo y CuantitativoDocumento8 páginasInforme Paradigma Cualitativo y CuantitativoRemy MendozaAún no hay calificaciones
- Vitaminas LiposolublesDocumento38 páginasVitaminas LiposolublesKaletAún no hay calificaciones
- Vitaminas y MineralesDocumento18 páginasVitaminas y MineralesGabriel Severiche ArancibiaAún no hay calificaciones
- Clase 1 Anatomia y FisiologiaDocumento26 páginasClase 1 Anatomia y FisiologiaRaquelita MoranAún no hay calificaciones
- Sistema LinfáticoDocumento18 páginasSistema LinfáticoFrancisca FuentevillaAún no hay calificaciones
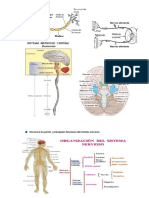
- Partes y Funciones Sistema NerviosoDocumento4 páginasPartes y Funciones Sistema Nerviosoicastillo1267Aún no hay calificaciones
- Sistema CardiovascularDocumento16 páginasSistema CardiovascularCARLOS MANUEL ELIAS PRINCIPE OSORIOAún no hay calificaciones
- Reconoce Las Partes y Principales Funciones Del Sistema NerviosoDocumento3 páginasReconoce Las Partes y Principales Funciones Del Sistema NerviososaloAún no hay calificaciones
- Osteologia DibujosDocumento11 páginasOsteologia Dibujosandrea gomezAún no hay calificaciones
- CelulaDocumento25 páginasCelulaEdgar TintaAún no hay calificaciones
- Vitaminas y MineralesDocumento6 páginasVitaminas y Mineralesamanda narvaez maciasAún no hay calificaciones
- Guía de Organelos CelularesDocumento6 páginasGuía de Organelos CelularesMaggie Tapia ZúñigaAún no hay calificaciones
- Calisifacion AlimenticiaDocumento5 páginasCalisifacion AlimenticiaMirnaDedeLeonAún no hay calificaciones
- Estructura Del ADNDocumento6 páginasEstructura Del ADNmillerlandy gutierrezAún no hay calificaciones
- Practica NDocumento5 páginasPractica NKarla NoriegaAún no hay calificaciones
- Metodo Cientifico)Documento24 páginasMetodo Cientifico)Luis fernando Cañola AcaroAún no hay calificaciones
- Sistema Nervioso Humano PDFDocumento8 páginasSistema Nervioso Humano PDFWilfredoNoleAguilarAún no hay calificaciones
- Alimentos FuncionalesDocumento27 páginasAlimentos FuncionalesLeonel EstradaAún no hay calificaciones
- Cuáles Son Las Funciones de Las ProteínasDocumento11 páginasCuáles Son Las Funciones de Las ProteínasCarlos CMAún no hay calificaciones
- MitosisDocumento7 páginasMitosisMarcos AntonioAún no hay calificaciones
- Ciclo Celular - Medicina 2022Documento16 páginasCiclo Celular - Medicina 2022Verduleria Los BajitosAún no hay calificaciones
- Introduccion A La Bioquimica en Nutricion 2021Documento88 páginasIntroduccion A La Bioquimica en Nutricion 2021Agus RomeroAún no hay calificaciones
- Diabetes 1Documento46 páginasDiabetes 1astrid carolina flores cabreraAún no hay calificaciones
- ENZIMASDocumento38 páginasENZIMASClaudia LoredoAún no hay calificaciones
- Sistema NerviosoDocumento51 páginasSistema NerviosoJuan Carlos Serrano TiradoAún no hay calificaciones
- Las Enzimas PDFDocumento9 páginasLas Enzimas PDFTriny VilladiegoAún no hay calificaciones
- BIOTECNOLOGIADocumento3 páginasBIOTECNOLOGIAdanielon14Aún no hay calificaciones
- Estructura y Función de Los Principales Tejidos en El OrganismoDocumento3 páginasEstructura y Función de Los Principales Tejidos en El OrganismoMayoTrujilloAún no hay calificaciones
- Tema 3 Metabolismo Lipidos 111 2 ParcialDocumento30 páginasTema 3 Metabolismo Lipidos 111 2 ParcialMario BownstherAún no hay calificaciones
- Sistema Linfatico (Histologia)Documento13 páginasSistema Linfatico (Histologia)Danielis BetancourtAún no hay calificaciones
- Clase No. 6 Estructura y Función de La CélulaDocumento53 páginasClase No. 6 Estructura y Función de La CélulaJoveth YanesAún no hay calificaciones
- Ensayo - Importancia de La Bioquímica en El Área de La Nutrición y La Dietética.Documento2 páginasEnsayo - Importancia de La Bioquímica en El Área de La Nutrición y La Dietética.jarithsa videsAún no hay calificaciones
- BioquimicaDocumento14 páginasBioquimica04144451342Aún no hay calificaciones
- Sistema GI Presentación Dra ChávezDocumento54 páginasSistema GI Presentación Dra ChávezMaría Antonieta ChávezAún no hay calificaciones
- Divicion CelularDocumento6 páginasDivicion CelularEna Margarita Mercado OlmosAún no hay calificaciones
- Moléculas Inorgánicas y OrgánicasDocumento5 páginasMoléculas Inorgánicas y Orgánicassilvana ArguelloAún no hay calificaciones
- 3 - VitaminasDocumento25 páginas3 - VitaminasGabriel ArambulaAún no hay calificaciones
- Manual de VitaminasDocumento41 páginasManual de VitaminasalexandraAún no hay calificaciones
- El Transporte CelularDocumento2 páginasEl Transporte CelularkeillapalaciosAún no hay calificaciones
- 8 Metabolismo de Los LipidosDocumento7 páginas8 Metabolismo de Los LipidosMervin Arauz HoyosAún no hay calificaciones
- Mapa Conceptual Del Sistema EndocrinoDocumento1 páginaMapa Conceptual Del Sistema EndocrinoKARLA PAOLA AGUIRRE CARRIÓNAún no hay calificaciones
- Organelos CelularesDocumento9 páginasOrganelos CelularesJuan Pablo MPAún no hay calificaciones
- Ind U1 A2 XxyzDocumento15 páginasInd U1 A2 XxyzArcelia PerezAún no hay calificaciones
- Funciones de Los Lipidos 2Documento20 páginasFunciones de Los Lipidos 2carmen santosAún no hay calificaciones
- Primera Clase de BiologiaDocumento28 páginasPrimera Clase de BiologiaLuxa Valverde ChicllaAún no hay calificaciones
- Clase Tema 6a GlucolisisDocumento53 páginasClase Tema 6a GlucolisisArianaRiosAún no hay calificaciones
- Carbohidratos ConejoDocumento5 páginasCarbohidratos Conejomonica100% (1)
- El Agua y Actividad Del Agua en Los Alimentos-2010Documento27 páginasEl Agua y Actividad Del Agua en Los Alimentos-2010Floral_Ana_Gut_6430Aún no hay calificaciones
- TejidosDocumento17 páginasTejidosIvanAún no hay calificaciones
- Los Organelos y EstructurasDocumento3 páginasLos Organelos y EstructurasMaría Alejandra Oyanadel GuerreroAún no hay calificaciones
- Anatomia y Fisiologia SistemasDocumento32 páginasAnatomia y Fisiologia SistemasSergio GalvanAún no hay calificaciones
- Act 24 BiologiaDocumento6 páginasAct 24 BiologiaItzel Guadalupe Lazcano VelázquezAún no hay calificaciones
- Miocinas y Regulación Metabólica, Una Revisión Sistemática: OriginalDocumento8 páginasMiocinas y Regulación Metabólica, Una Revisión Sistemática: OriginalRelviGuzmanApazaAún no hay calificaciones
- Hipopara Mas HipercalDocumento3 páginasHipopara Mas HipercalPaolo MessinaAún no hay calificaciones
- Alopecia en Antebrazos - Un Reto DiagnósticoDocumento5 páginasAlopecia en Antebrazos - Un Reto DiagnósticoPaolo MessinaAún no hay calificaciones
- Consenso de Prediabetes Documento de Posicion de La ALAD 2017Documento20 páginasConsenso de Prediabetes Documento de Posicion de La ALAD 2017Paolo MessinaAún no hay calificaciones
- Artículo - Riesgo de Malignidad de Los Nódulos Tiroideos Con Atipia de Significado InciertoDocumento4 páginasArtículo - Riesgo de Malignidad de Los Nódulos Tiroideos Con Atipia de Significado InciertoPaolo MessinaAún no hay calificaciones
- Artículo - Tratamiento Médico en El Síndrome de CushingDocumento4 páginasArtículo - Tratamiento Médico en El Síndrome de CushingPaolo MessinaAún no hay calificaciones
- Metabolismo de Proteínas y AminoácidosDocumento13 páginasMetabolismo de Proteínas y AminoácidosJuanAún no hay calificaciones
- Complejo Multienzimático Que Interviene en La Biosíntesis de Ácidos GrasosDocumento3 páginasComplejo Multienzimático Que Interviene en La Biosíntesis de Ácidos GrasosMichaelRvAún no hay calificaciones
- Plantas CamDocumento17 páginasPlantas Camanon_797932374Aún no hay calificaciones
- Complejo de La Piruvato DeshidrogenasaDocumento5 páginasComplejo de La Piruvato DeshidrogenasaMaria Zimbrelo AlcarriaAún no hay calificaciones
- Síntesis de Ácidos GrasosDocumento7 páginasSíntesis de Ácidos GrasosGCanisAún no hay calificaciones
- La Fermentacion y Sus TiposDocumento32 páginasLa Fermentacion y Sus TiposJuanPerez100% (1)
- Ciclo de La UreaDocumento47 páginasCiclo de La UreaSaid Figueroa DueñasAún no hay calificaciones
- Tipos de MetabolismoDocumento115 páginasTipos de MetabolismoJazive Alva100% (1)
- PreguntasDocumento4 páginasPreguntasgabrielaAún no hay calificaciones
- 10 Reacciones de La GlucolisisDocumento3 páginas10 Reacciones de La GlucolisisFernando CruzAún no hay calificaciones
- UROGENESISDocumento5 páginasUROGENESISRodrigo Acuña Leiva100% (1)
- PDF Sopa de Letras de Lipidos CompressxaDocumento1 páginaPDF Sopa de Letras de Lipidos CompressxaAlexis HianAún no hay calificaciones
- Glicólisis, Mecanismo de RegulaciónDocumento19 páginasGlicólisis, Mecanismo de RegulaciónJaneth SanchezAún no hay calificaciones
- Eduardo Díaz Balderas Felipe López Estudillo Jehur Mastranzo Osorio Jayr Zamora RojasDocumento27 páginasEduardo Díaz Balderas Felipe López Estudillo Jehur Mastranzo Osorio Jayr Zamora RojasJehur Mastranzo OsorioAún no hay calificaciones
- Activ 03 EdA 04 - 3° - 4° - Hoja ActividadesDocumento4 páginasActiv 03 EdA 04 - 3° - 4° - Hoja ActividadesJulio Cesar Ramos OrdoñezAún no hay calificaciones
- Cuestiones Autoevaluacion Metabolismo 1Documento2 páginasCuestiones Autoevaluacion Metabolismo 1Prof. Víctor M. VitoriaAún no hay calificaciones
- Infografía de Glucólisis.Documento1 páginaInfografía de Glucólisis.Ximena Cervantes LopezAún no hay calificaciones
- Metabolismo de LípidosDocumento6 páginasMetabolismo de LípidosKevinAlvaroMendozaAranibarAún no hay calificaciones
- Clase 2. Caracteristicas Biologicas de Los Seres VivosDocumento17 páginasClase 2. Caracteristicas Biologicas de Los Seres VivosPatricia LealAún no hay calificaciones
- Taller N 4 BioquimicaDocumento6 páginasTaller N 4 Bioquimicaadelina hernandezAún no hay calificaciones
- Gfdigestion de Las ProteinasDocumento4 páginasGfdigestion de Las ProteinasJoseph Martinez RamirezAún no hay calificaciones
- Bacterias QuimioautótrofasDocumento5 páginasBacterias QuimioautótrofasDoris Esther Asencio Cortez100% (1)
- Bio Cuestionario 6Documento3 páginasBio Cuestionario 6Lemon Tea86% (7)
- Practica 05 - Plantas C3 y C4 - Equipo 6Documento12 páginasPractica 05 - Plantas C3 y C4 - Equipo 6PL11PGAGONZALO HERNANDEZ AGUIRREAún no hay calificaciones
- Urea Casos ClinicosDocumento31 páginasUrea Casos ClinicosAnonymous XsmkCkHQAún no hay calificaciones
- Clase 9 - Reacciones Del Ciclo de Krebs y Balance Energético, Mecanismos de Regulación. Interacción Con Otras Rutas Metabólicas, Ciclo Del GlioxilatoDocumento29 páginasClase 9 - Reacciones Del Ciclo de Krebs y Balance Energético, Mecanismos de Regulación. Interacción Con Otras Rutas Metabólicas, Ciclo Del GlioxilatoAlex Ostos AlvaAún no hay calificaciones
- MicrobiologiaDocumento21 páginasMicrobiologiaEHdgar SalOrtegaAún no hay calificaciones
- Tejido AdiposoDocumento9 páginasTejido Adipososofia montenegroAún no hay calificaciones
- CEPRE Unsch PDFDocumento2 páginasCEPRE Unsch PDFjersonAún no hay calificaciones
- Cuestionario Integración de Carbohidratos I y IIDocumento3 páginasCuestionario Integración de Carbohidratos I y IIKedwin Montejo100% (1)