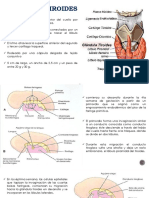
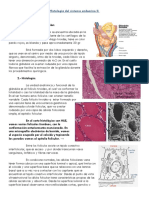
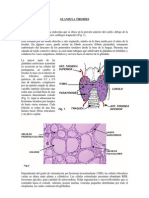
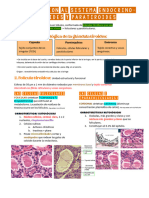
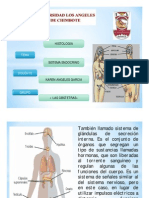
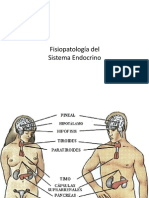
También podría gustarte
- Histología de La Glándula TiroidesDocumento5 páginasHistología de La Glándula TiroidesAnonymous W2RgWgucAún no hay calificaciones
- 11 Tiroides y ParatiroidesDocumento51 páginas11 Tiroides y ParatiroidesMariella Palacios SandovalAún no hay calificaciones
- 11 Tiroides y ParatiroidesDocumento51 páginas11 Tiroides y ParatiroidesFARIDE DANIELA BOGGIO CUENCAAún no hay calificaciones
- Tiroides Estructura y Funcion ModificadoDocumento50 páginasTiroides Estructura y Funcion ModificadoCarmen Juscamaita VegaAún no hay calificaciones
- Glándula Tiroides 1Documento12 páginasGlándula Tiroides 1Fede AranaAún no hay calificaciones
- Informe de TiroidesDocumento6 páginasInforme de TiroidesIngrid Andreina Castillo VizcayaAún no hay calificaciones
- HistologiaDocumento12 páginasHistologiaJodie LeónAún no hay calificaciones
- Histologia Del Sistema Endocrino IIDocumento12 páginasHistologia Del Sistema Endocrino IIPedro GonzalezAún no hay calificaciones
- Histología de La Glándula TiroidesDocumento6 páginasHistología de La Glándula TiroidesLucia Saavedra LozadaAún no hay calificaciones
- Histologia Del Sistema Endocrino Piero Koo Doig PDFDocumento5 páginasHistologia Del Sistema Endocrino Piero Koo Doig PDFLuis Carlos Calvanapon TerronesAún no hay calificaciones
- Tiro IdesDocumento3 páginasTiro IdesAlfredo OyarzunAún no hay calificaciones
- Sistema Endocrino IIDocumento42 páginasSistema Endocrino IIsuse santosAún no hay calificaciones
- Tema 37Documento20 páginasTema 37sarahyAún no hay calificaciones
- Glándula Tiroidea HistologiaDocumento5 páginasGlándula Tiroidea HistologiaMaría Alejandra Aznate HernándezAún no hay calificaciones
- Histologia Del Aparato Reproductor MasculinoDocumento5 páginasHistologia Del Aparato Reproductor MasculinoMauricio Leon100% (1)
- Scheznarda EndocrinoDocumento7 páginasScheznarda Endocrinoricardo_roldan_36Aún no hay calificaciones
- Histologia Ii - MC SRMDocumento8 páginasHistologia Ii - MC SRMAmarildo CristinoAún no hay calificaciones
- Tejidos Del Sistema Endocrino - Sheila MendozaDocumento11 páginasTejidos Del Sistema Endocrino - Sheila MendozaShei MendozaAún no hay calificaciones
- Resumen de TiroidesDocumento6 páginasResumen de TiroidesGuille HerreraAún no hay calificaciones
- Tiroides PDFDocumento21 páginasTiroides PDFJosue Mendoza MárquezAún no hay calificaciones
- Apuntes EndocrinoDocumento197 páginasApuntes EndocrinoMishel VillaAún no hay calificaciones
- Glandula TiroidesDocumento6 páginasGlandula TiroidesNikole Samantha Castro MartínezAún no hay calificaciones
- Tiroides ParatiroidesDocumento4 páginasTiroides Paratiroidesestefani.ortiz.pAún no hay calificaciones
- CloroplastosDocumento4 páginasCloroplastosErnesto RomanAún no hay calificaciones
- Anatomía y Fisiología de La Glándula TiroidesDocumento4 páginasAnatomía y Fisiología de La Glándula TiroidesLeslie Thalia Rojas JustinianoAún no hay calificaciones
- Aparato Reproductor FemeninoDocumento11 páginasAparato Reproductor FemeninoflorenciarenieroAún no hay calificaciones
- Glandulas Suprarrenaes 2Documento5 páginasGlandulas Suprarrenaes 2Amarildo CristinoAún no hay calificaciones
- TIROIDESDocumento16 páginasTIROIDESdanielaAún no hay calificaciones
- Glándulas SuprarrenalesDocumento27 páginasGlándulas SuprarrenalesMarianela EscobarAún no hay calificaciones
- Fisiología de La Gládula TiroidesDocumento19 páginasFisiología de La Gládula TiroidesAngie SaileAún no hay calificaciones
- OVARIODocumento4 páginasOVARIOPedro GuillenAún no hay calificaciones
- 140 - Fisiología de Las Glándulas Tiroides y ParatiroidesDocumento18 páginas140 - Fisiología de Las Glándulas Tiroides y ParatiroidesAnita AlanizAún no hay calificaciones
- 2 Monografia de FisiopatoDocumento17 páginas2 Monografia de FisiopatoANABEL ZARATE CUSIAún no hay calificaciones
- Aparato Reproductor FemeninoDocumento10 páginasAparato Reproductor FemeninoMANRIQUE GUTIÉRREZ STEPHANY GIOMAYRAAún no hay calificaciones
- Resumen de CloroplastosDocumento4 páginasResumen de Cloroplastospaz_saavedra_350% (2)
- HISTOLOGIA DEL SISTEMA REPRODUCTOR MASCULINO y FemeninoDocumento17 páginasHISTOLOGIA DEL SISTEMA REPRODUCTOR MASCULINO y FemeninoEstuardo Len100% (1)
- 14 Reproductor FemeninoDocumento90 páginas14 Reproductor FemeninoPaty AlamoAún no hay calificaciones
- Clase 2-Biología Gen 24 VeranoDocumento13 páginasClase 2-Biología Gen 24 Veranoacostajohn397Aún no hay calificaciones
- Sist. Rep. FemeninoDocumento69 páginasSist. Rep. FemeninoUSMP FN ARCHIVOSAún no hay calificaciones
- Exposición Sobre El Sistema EndocrinoDocumento19 páginasExposición Sobre El Sistema EndocrinoNancy Yurisel Gonzales FloresAún no hay calificaciones
- CUESTIONARIO 14 Ramírez CiezaDocumento10 páginasCUESTIONARIO 14 Ramírez CiezaDIANA RAMIREZ CIEZAAún no hay calificaciones
- Histologia ManualDocumento127 páginasHistologia ManualDinora Alejandra E MAún no hay calificaciones
- Glándula TiroidesDocumento3 páginasGlándula TiroidesChucho MéndezAún no hay calificaciones
- Fisiologia TiroidesDocumento27 páginasFisiologia TiroidesGabrielaAún no hay calificaciones
- Tema 1 - Tiroides, Hipotiroidismo y Coma Mixedematoso.Documento14 páginasTema 1 - Tiroides, Hipotiroidismo y Coma Mixedematoso.Medicina 5 Seccion 11Aún no hay calificaciones
- Tiroides y Paratiroides - HistologiaDocumento21 páginasTiroides y Paratiroides - HistologiagorditobondiolauwuAún no hay calificaciones
- Hormonas TiroideasDocumento17 páginasHormonas TiroideasMonserrat Blanquel García100% (1)
- Sistema de Membranas CitoplasmicasDocumento5 páginasSistema de Membranas CitoplasmicasToloza VladimirAún no hay calificaciones
- Organelas 20citoplasmaticas 2013Documento17 páginasOrganelas 20citoplasmaticas 2013Luilly FernandoAún no hay calificaciones
- Aparato Reproductor FemeninoDocumento77 páginasAparato Reproductor FemeninoUSMP FN ARCHIVOS100% (1)
- Glandula TiroidesDocumento33 páginasGlandula TiroidesNayelli Farías.100% (1)
- Reticulo Endoplasmatico LisoDocumento7 páginasReticulo Endoplasmatico Lisomarlen cruz jaureguiAún no hay calificaciones
- Glandulas SuprarrenalesDocumento9 páginasGlandulas SuprarrenalesJosselyn LoorAún no hay calificaciones
- Clase 7 ConjuntivoDocumento2 páginasClase 7 ConjuntivoEmili Wiemberg FredesAún no hay calificaciones
- TIROIDESDocumento26 páginasTIROIDEScumbajindanielaAún no hay calificaciones
- FisioexamenDocumento15 páginasFisioexamenMarié AguilarAún no hay calificaciones
- Tejido HematopoyeticoDocumento78 páginasTejido HematopoyeticoAlba_luna27Aún no hay calificaciones
- Anemia Para Todos: Medicina Para TodosDe EverandAnemia Para Todos: Medicina Para TodosCalificación: 3 de 5 estrellas3/5 (1)
- Coagulación Para Todos: Medicina Para TodosDe EverandCoagulación Para Todos: Medicina Para TodosCalificación: 4.5 de 5 estrellas4.5/5 (3)
- DocumentoDocumento2 páginasDocumentoluisAún no hay calificaciones
- Repaso El Examen Físico Normal de TóraxDocumento4 páginasRepaso El Examen Físico Normal de TóraxluisAún no hay calificaciones
- Exposición ColonDocumento3 páginasExposición ColonluisAún no hay calificaciones
- Indicadores en SaludDocumento4 páginasIndicadores en Saludluis100% (1)
- Autoexploración y Autoconocimiento de Las MamasDocumento12 páginasAutoexploración y Autoconocimiento de Las MamasluisAún no hay calificaciones
- Fichas Boa 2Documento3 páginasFichas Boa 2luisAún no hay calificaciones
- Fichas Boa 3Documento6 páginasFichas Boa 3luisAún no hay calificaciones
- Fichas Boa 4Documento5 páginasFichas Boa 4luisAún no hay calificaciones
- Trabajo Grupo 3Documento11 páginasTrabajo Grupo 3luisAún no hay calificaciones
- GonorreaDocumento2 páginasGonorrealuisAún no hay calificaciones
- Guia para IntegradorDocumento10 páginasGuia para IntegradorluisAún no hay calificaciones
- Describa La Anatomía Del Cordón EspermáticoDocumento3 páginasDescriba La Anatomía Del Cordón EspermáticoluisAún no hay calificaciones
- Glandulas Sexuales AccesoriasDocumento4 páginasGlandulas Sexuales AccesoriasluisAún no hay calificaciones
- Las EmocionesDocumento13 páginasLas Emocionesyuviskar urdanetaAún no hay calificaciones
- Dra Rosa NovoaDocumento24 páginasDra Rosa NovoarocioAún no hay calificaciones
- Emergencias y Urgencias EndocrinológicasDocumento47 páginasEmergencias y Urgencias EndocrinológicasJhuliana Sanchez ChocanoAún no hay calificaciones
- Sistema Endocrino y Las HormonasDocumento3 páginasSistema Endocrino y Las HormonasJorge Vilela CespedesAún no hay calificaciones
- Sistema EndocrinoDocumento19 páginasSistema EndocrinoHeber FallenAún no hay calificaciones
- EndocrinopatiasDocumento94 páginasEndocrinopatiastititoledoAún no hay calificaciones
- Fisiologia SuprarrenalDocumento36 páginasFisiologia SuprarrenalH Vásquez Galindo100% (2)
- Funcion SuprarrenalDocumento5 páginasFuncion SuprarrenalQUIMICO CLINICO WILLIANS SANCHEZ100% (1)
- HORMONASDocumento25 páginasHORMONASLeydi Yudith Angarita Bautista100% (22)
- Tabla de EndocrinologiaDocumento3 páginasTabla de EndocrinologiaFroi BalboaAún no hay calificaciones
- Neurociencia AfectivaDocumento5 páginasNeurociencia AfectivaXimena OrtegaAún no hay calificaciones
- Parte de Endocrino RemelDocumento3 páginasParte de Endocrino RemelRemel Mendieta EspinozaAún no hay calificaciones
- Hiperplasia Suprarrenal CongénitaDocumento4 páginasHiperplasia Suprarrenal CongénitaVicente Parada AguirreAún no hay calificaciones
- 8 Presentacion Glandulas Endocrinas 230407Documento33 páginas8 Presentacion Glandulas Endocrinas 230407Anabelitha FloresAún no hay calificaciones
- Sindrome de La Tiroides 1Documento232 páginasSindrome de La Tiroides 1Yddemar BrownAún no hay calificaciones
- CAPITULO I Ansiedad-1Documento12 páginasCAPITULO I Ansiedad-1Marco GarridoAún no hay calificaciones
- GLANDULASDocumento3 páginasGLANDULASJody EcAún no hay calificaciones
- Generalidades, Partes y Funciones Del Sistema EndocrinoDocumento13 páginasGeneralidades, Partes y Funciones Del Sistema EndocrinoKaren Alejandra CERQUERA TRUJILLOAún no hay calificaciones
- AdrenocorticalesDocumento10 páginasAdrenocorticalesClaudia Ana Sotomayor MolinaAún no hay calificaciones
- Fisiopatología Estereidogenesis SuprarreanalDocumento5 páginasFisiopatología Estereidogenesis SuprarreanalFrancisco DavidAún no hay calificaciones
- 07 Endocrinología 2da Fase PDFDocumento61 páginas07 Endocrinología 2da Fase PDFJessenia JudithAún no hay calificaciones
- Enfermedad de AddisonDocumento5 páginasEnfermedad de Addisonkarelys jimenezAún no hay calificaciones
- Sistema EndocrinoDocumento37 páginasSistema EndocrinoMaurilio RodriguezAún no hay calificaciones
- La Cura de La Fatiga AdrenalDocumento7 páginasLa Cura de La Fatiga AdrenalMiguel Gutiérrez Márquez100% (1)
- La PielDocumento21 páginasLa PielanahiAún no hay calificaciones
- Articulo de La Dra PHD Marianela Castés Médico InmunólogoDocumento4 páginasArticulo de La Dra PHD Marianela Castés Médico Inmunólogo14057022Aún no hay calificaciones
- Neurobiología Del Buen TratoDocumento39 páginasNeurobiología Del Buen TratoFelipe Aravena100% (1)
- Envejecimiento Del Sistema Endocrino 2.Documento63 páginasEnvejecimiento Del Sistema Endocrino 2.Roley EspinosaAún no hay calificaciones
- Medicina y Laboratorio en Psiquiatria y Ciencias de La ConductaDocumento443 páginasMedicina y Laboratorio en Psiquiatria y Ciencias de La Conductazsmm1993Aún no hay calificaciones
- Recomendaciones SOCHIMI Sepsis. FinalDocumento13 páginasRecomendaciones SOCHIMI Sepsis. FinalJavieraAlondraDiazAún no hay calificaciones