También podría gustarte
- Introducción a la Biología: RESÚMENES UNIVERSITARIOSDe EverandIntroducción a la Biología: RESÚMENES UNIVERSITARIOSCalificación: 5 de 5 estrellas5/5 (1)
- Entrevista 1Documento9 páginasEntrevista 1Works DCAún no hay calificaciones
- Taller EntrecruzamientoDocumento6 páginasTaller EntrecruzamientoMiriam CaroAún no hay calificaciones
- Modulo Ingles PDFDocumento88 páginasModulo Ingles PDFBrenda Lara RuizAún no hay calificaciones
- Manejo de TB Consenso MéxicanoDocumento116 páginasManejo de TB Consenso MéxicanoAdriana Asturizaga100% (1)
- Ligamiento, EntrecruzamientoDocumento33 páginasLigamiento, EntrecruzamientoCUCBA Biologia100% (8)
- Informe de Aislamiento de HongosDocumento29 páginasInforme de Aislamiento de HongosJesid Angel Ticse Huamán50% (4)
- Introduccion A La GeneticaDocumento36 páginasIntroduccion A La GeneticaAlejandro Mansilla RodriguezAún no hay calificaciones
- Cromosoma GENERALIDADESDocumento11 páginasCromosoma GENERALIDADESMaryuriVegaAún no hay calificaciones
- Psicobiología Tema 2 ResumenDocumento11 páginasPsicobiología Tema 2 ResumenArabiaDíazAún no hay calificaciones
- ClaseDocumento9 páginasClaseAnonymous zsSgnZj2Aún no hay calificaciones
- Preguntas y Respuestas GenéticaDocumento10 páginasPreguntas y Respuestas Genéticarega0683% (6)
- Taller #5Documento10 páginasTaller #5kmilo159Aún no hay calificaciones
- Genetica Teoriacromosmica Bio2clase2011bDocumento69 páginasGenetica Teoriacromosmica Bio2clase2011bOscar Trujillo AcevedoAún no hay calificaciones
- CromosomaDocumento14 páginasCromosomaSarahi SilvaAún no hay calificaciones
- Resumen Segundo ParcialDocumento13 páginasResumen Segundo ParcialRocio DabosAún no hay calificaciones
- Apuntes Tema 3Documento25 páginasApuntes Tema 3luciavs25Aún no hay calificaciones
- PolihibridoDocumento30 páginasPolihibridoCharlin Cerna RuizAún no hay calificaciones
- Apuntes de Clase BiocelDocumento5 páginasApuntes de Clase BiocelConsuelo Kathaline Gallardo MondacaAún no hay calificaciones
- GENÉTICADocumento18 páginasGENÉTICAGustavo HernándezAún no hay calificaciones
- Genética PostmendelianaDocumento9 páginasGenética PostmendelianaVentitasSantiagoAún no hay calificaciones
- Resumen de GenéticaDocumento5 páginasResumen de GenéticaJogelina zuñigaAún no hay calificaciones
- T3 Genética MendelianaDocumento11 páginasT3 Genética MendelianaÁlvaro Pereda EscuderoAún no hay calificaciones
- Teoría CromosómicaDocumento20 páginasTeoría CromosómicakevinAún no hay calificaciones
- 17-Ligamiento y Recombinación en EucariontesDocumento25 páginas17-Ligamiento y Recombinación en EucariontesAlmudena MeiAún no hay calificaciones
- CromosomasDocumento8 páginasCromosomasMiguel Angel Rodas HerreraAún no hay calificaciones
- UNIDAD 2 GENETICA MEDICA 3ra PARTEDocumento49 páginasUNIDAD 2 GENETICA MEDICA 3ra PARTEAndrea RieraAún no hay calificaciones
- Tema 5 GmaDocumento19 páginasTema 5 GmaanabelenorcajadapastorAún no hay calificaciones
- GeneticaDocumento32 páginasGeneticaoriana pintoAún no hay calificaciones
- Clase 11 División de Células SexualesDocumento36 páginasClase 11 División de Células SexualesJoel Pinedo PacayaAún no hay calificaciones
- Biología GenéticaDocumento106 páginasBiología Genéticajassy graciaAún no hay calificaciones
- Concepto Moderno de ObstetriciaDocumento19 páginasConcepto Moderno de ObstetriciaPriscilla Sandoya Bernabé100% (1)
- UNIDAD 3 BIOLOGIA UNM Teoricas 2023Documento102 páginasUNIDAD 3 BIOLOGIA UNM Teoricas 2023Sintia RedAún no hay calificaciones
- 5 - Genetica y ReplicacionDocumento29 páginas5 - Genetica y Replicacionjose.torres1Aún no hay calificaciones
- Cromosomas Sexuales.Documento25 páginasCromosomas Sexuales.Karol cruzAún no hay calificaciones
- Apuntes Unidad 4Documento18 páginasApuntes Unidad 4Karla Edith CortesAún no hay calificaciones
- CromosomasDocumento12 páginasCromosomasCarolina MarteAún no hay calificaciones
- Capitulo 7 KlugDocumento10 páginasCapitulo 7 KlugIngrid Anaya MoralesAún no hay calificaciones
- Genetica Ligamiento EntrecruzamientoDocumento17 páginasGenetica Ligamiento EntrecruzamientodbzoAún no hay calificaciones
- Leyes de Mendel ImportanciaDocumento5 páginasLeyes de Mendel ImportanciaKAREN SOFIA RAMIREZ SOACHEAún no hay calificaciones
- Terminología GenéticaDocumento6 páginasTerminología GenéticaKeviin GomëZz K-o0% (1)
- Reproducción Sexual y Meiosis - Unidad 10 Parte 2Documento49 páginasReproducción Sexual y Meiosis - Unidad 10 Parte 2Jaqueline Alexias LozanoAún no hay calificaciones
- Monografía Foro 8Documento24 páginasMonografía Foro 8cesar davidAún no hay calificaciones
- Genética Mendeliana 1Documento44 páginasGenética Mendeliana 1Germain Rider FigueroaAún no hay calificaciones
- Taller BiologiaDocumento20 páginasTaller BiologiaCristhian Andres Rojas GrisalesAún no hay calificaciones
- Gentica 1Documento54 páginasGentica 1ELSY HUARACHI PALOMINOAún no hay calificaciones
- CROMOSOMASDocumento4 páginasCROMOSOMASCarlos Rivera JaraAún no hay calificaciones
- Biologia - Semana 3 - Tema2Documento10 páginasBiologia - Semana 3 - Tema2Marilyn StefaniaAún no hay calificaciones
- GM - Guía de Estudio #4Documento12 páginasGM - Guía de Estudio #4mczanaboni100% (2)
- 12.wuolah Premium Tema XIIDocumento5 páginas12.wuolah Premium Tema XII2mn2kzd4h2Aún no hay calificaciones
- Taller EntrecruzamientoDocumento6 páginasTaller EntrecruzamientoYARITZA VANESSA BARCELO OJEDAAún no hay calificaciones
- 25 CromosomasDocumento9 páginas25 CromosomasshiyrrAún no hay calificaciones
- Estructura de Los CromosomasDocumento12 páginasEstructura de Los CromosomasRosa CorderoAún no hay calificaciones
- Genetica Completo FinalDocumento75 páginasGenetica Completo FinalPablo AparicioAún no hay calificaciones
- Proyecto de GenomaDocumento16 páginasProyecto de GenomaadrianaAún no hay calificaciones
- Clase de Rasgos MonogenicosDocumento81 páginasClase de Rasgos MonogenicosCapry De Leon RadaAún no hay calificaciones
- Informe Teoria Cromosomica Andres EscobarDocumento4 páginasInforme Teoria Cromosomica Andres EscobarAndrés EscobarAún no hay calificaciones
- Genética No MendelianaDocumento28 páginasGenética No MendelianaAlexander Oviedo BenitezAún no hay calificaciones
- BIOTEXTODocumento12 páginasBIOTEXTOLina Luz Perez MedranoAún no hay calificaciones
- Unidad 3 Patologia Unefm - DR Alvaro E. Ordaz R.Documento44 páginasUnidad 3 Patologia Unefm - DR Alvaro E. Ordaz R.achraf abboudAún no hay calificaciones
- BiologiaDocumento20 páginasBiologiaRoberto CastellanosAún no hay calificaciones
- 3.2 CromosomasDocumento22 páginas3.2 CromosomasDaniel JaramilloAún no hay calificaciones
- Ligamiento y Mapeo CromosómicoDocumento7 páginasLigamiento y Mapeo CromosómicoAnisa Achhodi DadabhaiAún no hay calificaciones
- La Herencia BiologicaDocumento80 páginasLa Herencia Biologicakaren crisol solis linoAún no hay calificaciones
- Apuntes MirDocumento47 páginasApuntes MirRo11Aún no hay calificaciones
- EMBRIOLOGIADocumento5 páginasEMBRIOLOGIARo11Aún no hay calificaciones
- BIOESTADISTICADocumento4 páginasBIOESTADISTICARo11Aún no hay calificaciones
- HISTOLOGIADocumento4 páginasHISTOLOGIARo11Aún no hay calificaciones
- Acalasia 341832 Downloable 1693576Documento4 páginasAcalasia 341832 Downloable 1693576Ro11Aún no hay calificaciones
- Apuntes Mir2Documento5 páginasApuntes Mir2Ro11Aún no hay calificaciones
- MICROBIOLOGIADocumento5 páginasMICROBIOLOGIARo11Aún no hay calificaciones
- FISIOLOGIADocumento9 páginasFISIOLOGIARo11Aún no hay calificaciones
- CITOLOGIADocumento3 páginasCITOLOGIARo11Aún no hay calificaciones
- Met AeronauticaDocumento17 páginasMet Aeronauticagonzalo gorositoAún no hay calificaciones
- BIOQUIMICADocumento7 páginasBIOQUIMICARo11Aún no hay calificaciones
- Horario SemanalDocumento1 páginaHorario SemanalRo11Aún no hay calificaciones
- Indice MicroDocumento1 páginaIndice MicroRo11Aún no hay calificaciones
- Meteorología Aeronáutica Font TullotDocumento5 páginasMeteorología Aeronáutica Font TullotRo11Aún no hay calificaciones
- Apuntes MeteorologiaDocumento12 páginasApuntes MeteorologiaAdrian GuzmanAún no hay calificaciones
- Calendario 2020Documento12 páginasCalendario 2020Ro11Aún no hay calificaciones
- Calendario Completo de LaLiga SantanderDocumento13 páginasCalendario Completo de LaLiga SantanderLa VanguardiaAún no hay calificaciones
- Calendario LigaDocumento15 páginasCalendario Ligaismaeldeus8204Aún no hay calificaciones
- Calendario Liga 2019Documento5 páginasCalendario Liga 2019Ro11Aún no hay calificaciones
- Calendario Semanal 2015Documento3 páginasCalendario Semanal 2015scribdbAún no hay calificaciones
- Tecnicas Utilizadas en Histología y MicrosDocumento32 páginasTecnicas Utilizadas en Histología y MicrosHurtado MafeAún no hay calificaciones
- Ecologia de Las ComunidadesDocumento12 páginasEcologia de Las ComunidadesHuereqq LambayeqAún no hay calificaciones
- AmortiguadoresDocumento4 páginasAmortiguadoresrocio_11Aún no hay calificaciones
- Taller Ciclo Celular SeptimoDocumento2 páginasTaller Ciclo Celular SeptimoJOHNY MORALESAún no hay calificaciones
- Juana Cordova - A La OrillaDocumento51 páginasJuana Cordova - A La OrillaAndres MendezAún no hay calificaciones
- Neodent WEB PDFDocumento109 páginasNeodent WEB PDFIgnacio Aliaga ArellanoAún no hay calificaciones
- Fitoestrógenos en La Alimentación AnimalDocumento7 páginasFitoestrógenos en La Alimentación AnimalJean Piero L.EAún no hay calificaciones
- Lobo Ártico o Lobo de NieveDocumento9 páginasLobo Ártico o Lobo de NieveOdimar SayagoAún no hay calificaciones
- Proyecto Nutricion DiazDocumento16 páginasProyecto Nutricion DiazOslercap Caup PicotAún no hay calificaciones
- Informe N5 QB - Actividad Enzimatica de La Beta-GalactosidasaDocumento7 páginasInforme N5 QB - Actividad Enzimatica de La Beta-GalactosidasaKata VenegasAún no hay calificaciones
- Evaluación Diagnóstica de Biologia 1 para Tercer SemestreDocumento8 páginasEvaluación Diagnóstica de Biologia 1 para Tercer SemestreGustavo Adolfo García CollmanAún no hay calificaciones
- Reporte Práctica Biología - 3Documento7 páginasReporte Práctica Biología - 3Sara AguirreAún no hay calificaciones
- Semana 04 - S4 Sistema Piramidal y ExtrapiramidalDocumento61 páginasSemana 04 - S4 Sistema Piramidal y ExtrapiramidalAlonso L:3Aún no hay calificaciones
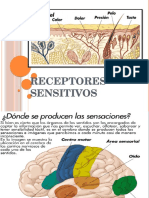
- Receptores SensitivosDocumento14 páginasReceptores SensitivostiareAún no hay calificaciones
- Psicologia Evolutiva Enfermeria Ii Semestre InvestigacionDocumento7 páginasPsicologia Evolutiva Enfermeria Ii Semestre Investigacionyury sanchezAún no hay calificaciones
- Vocabulario de BiologiaDocumento8 páginasVocabulario de BiologiaHernán MârtinezAún no hay calificaciones
- 05 Herramienta Corte PDFDocumento69 páginas05 Herramienta Corte PDFNevsaint MartinezAún no hay calificaciones
- Darwin, Genética y BiodiversidadDocumento10 páginasDarwin, Genética y BiodiversidadDavid VillaAún no hay calificaciones
- 22-Bases de Datos-BioinformDocumento21 páginas22-Bases de Datos-Bioinformmateo alvaradoAún no hay calificaciones
- 10) Espacio Visceral de Cuello 2018Documento10 páginas10) Espacio Visceral de Cuello 2018Catalina Ruiz LópezAún no hay calificaciones
- Dermatosis Perforantes - UpToDateDocumento57 páginasDermatosis Perforantes - UpToDateAnghelino Jesus Meza CentenoAún no hay calificaciones
- EF - III - Iniciativa y Toma de Decisiones - Mi Círculo de Influencia - V.1.2Documento23 páginasEF - III - Iniciativa y Toma de Decisiones - Mi Círculo de Influencia - V.1.2Benitez Marcos AtibenaAún no hay calificaciones
- Info08 PigmentosDocumento12 páginasInfo08 PigmentosSamuel Cruz Guillén100% (1)
- Programación 2017-2 OrienteDocumento17 páginasProgramación 2017-2 Orientedavid duqueAún no hay calificaciones
- Anatomiadetorax 190306023212Documento18 páginasAnatomiadetorax 190306023212Jimenez JaimeAún no hay calificaciones
- 51 - Emprender - Unidad 1 (Pag10-30)Documento21 páginas51 - Emprender - Unidad 1 (Pag10-30)florAún no hay calificaciones
- Tasa Metabolica Iso 8996.Documento14 páginasTasa Metabolica Iso 8996.danmolar100% (2)