También podría gustarte
- Histología Digestivo y AnexosDocumento29 páginasHistología Digestivo y AnexosJaviera Paz Guerrero CassanelloAún no hay calificaciones
- Tejido ConectivoDocumento18 páginasTejido Conectivo440sobreterrenoAún no hay calificaciones
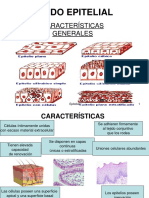
- 3.-Tejido EpitelialDocumento24 páginas3.-Tejido EpitelialAvassa100% (2)
- Preguntas Capitulo 4Documento6 páginasPreguntas Capitulo 4fredys galindoAún no hay calificaciones
- Cuestionario Tej. EpitelialDocumento8 páginasCuestionario Tej. EpitelialMónica González100% (2)
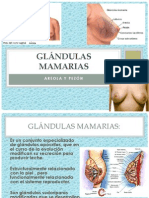
- Glándulas Mamarias PresentacionDocumento33 páginasGlándulas Mamarias PresentacionKarla A. Castro100% (1)
- Fichas Tejido ConectivoDocumento23 páginasFichas Tejido ConectivoMicaela HerrAún no hay calificaciones
- Piaget Vs Vygotsky.... para El Foro Academico de Anlly LuzDocumento4 páginasPiaget Vs Vygotsky.... para El Foro Academico de Anlly Luzluis grabielAún no hay calificaciones
- Aparato DigestivoDocumento37 páginasAparato DigestivoIvonne Rodriguez100% (3)
- Resumen Cap. 1 GuytonDocumento2 páginasResumen Cap. 1 GuytonMaité EspejoAún no hay calificaciones
- Integración de La Economía Circular en La Estrategia de SostenibilidadDocumento70 páginasIntegración de La Economía Circular en La Estrategia de SostenibilidadComunicarSe-Archivo100% (2)
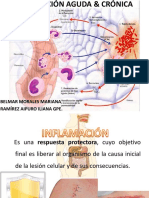
- Inflamación Aguda & CrónicaDocumento50 páginasInflamación Aguda & CrónicaMariana Belmar100% (1)
- Ojo Histologia Ross PDFDocumento14 páginasOjo Histologia Ross PDFReynis Monción33% (3)
- Tejido MuscularDocumento5 páginasTejido Muscularkatherine zambrano100% (1)
- Tejido ConectivoDocumento18 páginasTejido Conectivoerikparedes100% (4)
- Bioseguridad en El AnfiteatroDocumento16 páginasBioseguridad en El AnfiteatroPavel Luna50% (2)
- Marc Bloch - La Sociedad FeudalDocumento528 páginasMarc Bloch - La Sociedad FeudalEnrique Luque García100% (4)
- Fisiología de Guyton Resumen Capítulo 2Documento2 páginasFisiología de Guyton Resumen Capítulo 2Pablo Andrés Zurita93% (15)
- Clase Histologia. Aparato Digestivo IiDocumento65 páginasClase Histologia. Aparato Digestivo Iideimos100% (2)
- Tejido ConectivoDocumento23 páginasTejido ConectivoDanielaRojasAún no hay calificaciones
- Guía Mundo Descabellado 1Documento3 páginasGuía Mundo Descabellado 1Bárbara Madariaga González100% (1)
- Sistema Hematopoyético (Autoguardado)Documento30 páginasSistema Hematopoyético (Autoguardado)diana zanzAún no hay calificaciones
- Diapositivas Tejido Epitelial - PPTX MejoradasDocumento80 páginasDiapositivas Tejido Epitelial - PPTX MejoradasAdy Bedoya75% (4)
- Tejido MuscularDocumento5 páginasTejido MuscularKatiAriasAún no hay calificaciones
- Renovación, Reparación y Regeneración TisularDocumento57 páginasRenovación, Reparación y Regeneración TisularJUANITA ANDREA VALDIVIA JIMENEZ100% (1)
- Aparato UrinarioDocumento7 páginasAparato Urinariocarlos noriega diazAún no hay calificaciones
- Trafico Intracelular-SulyDocumento3 páginasTrafico Intracelular-SulySuly VigoAún no hay calificaciones
- LA PIEL, Generalidades 2020 PDFDocumento11 páginasLA PIEL, Generalidades 2020 PDFHuang Shan LinAún no hay calificaciones
- Ross Capitulo 15Documento12 páginasRoss Capitulo 15Ximena Ramirez GarciaAún no hay calificaciones
- Tejido Epitelial PDFDocumento61 páginasTejido Epitelial PDFSantiago MayaAún no hay calificaciones
- Teoria Tejido EpitelialDocumento74 páginasTeoria Tejido Epitelialstephanie reyes ruizAún no hay calificaciones
- Vías EndocíticasDocumento4 páginasVías EndocíticasSOFIA HAún no hay calificaciones
- 8 PleuraDocumento26 páginas8 PleuraNickyRbAún no hay calificaciones
- Capitulo 7 - Langman - Cavidades CorporalesDocumento3 páginasCapitulo 7 - Langman - Cavidades Corporalesmaria jose sanchez gilAún no hay calificaciones
- GLOSARIO TeguDocumento2 páginasGLOSARIO TeguZztrelliita WonkqAa LugooAún no hay calificaciones
- MORFOLOGIA Tejido EpitelialDocumento10 páginasMORFOLOGIA Tejido EpitelialcoroAún no hay calificaciones
- AXILADocumento41 páginasAXILAKiara SenmacheAún no hay calificaciones
- Señalizacion YuxtacrinaDocumento1 páginaSeñalizacion YuxtacrinaAlexanther Palomino Reategui100% (1)
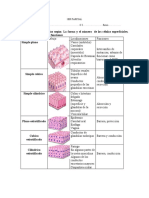
- Cuadro de EpiteliosDocumento5 páginasCuadro de EpiteliosMaría AlbornozAún no hay calificaciones
- (EV) S.digestivo III, GlandulasDocumento10 páginas(EV) S.digestivo III, GlandulasareliAún no hay calificaciones
- DGB y DGT PresentaciónDocumento36 páginasDGB y DGT PresentaciónEdgaris Sc'r0% (1)
- Escroto y Envolturas Del Testículo y Del EpidídimoDocumento4 páginasEscroto y Envolturas Del Testículo y Del EpidídimoSamantha RodriguezAún no hay calificaciones
- Tejido ConectivoDocumento54 páginasTejido ConectivoMichelle MGAún no hay calificaciones
- Tejido EpitelialDocumento4 páginasTejido EpitelialanaAún no hay calificaciones
- Mesodermo y MesenquimaDocumento3 páginasMesodermo y MesenquimaAntoAlvaradoAún no hay calificaciones
- Taller 2 PDFDocumento3 páginasTaller 2 PDFGénesis LeónAún no hay calificaciones
- Preguntas Primera Semana EmbriologiaDocumento1 páginaPreguntas Primera Semana Embriologiahumberto100% (2)
- Embriologia Del Sistema DigestivoDocumento45 páginasEmbriologia Del Sistema DigestivoJcaminoAún no hay calificaciones
- Membrana BasalDocumento14 páginasMembrana BasalJESSYRDGZAún no hay calificaciones
- Disco Germinativo TrilaminarDocumento64 páginasDisco Germinativo TrilaminarLorena JiménezAún no hay calificaciones
- Ross - Sistema CardiovascularDocumento2 páginasRoss - Sistema Cardiovascularijorgek75% (4)
- Monografia de Ramon Tejido ConectivoDocumento15 páginasMonografia de Ramon Tejido ConectivoJose Antonio Rivero Velásquez50% (2)
- Membrana BasalDocumento4 páginasMembrana BasalGleyser Jimmy100% (2)
- GametosDocumento4 páginasGametosGrisel RodriguezAún no hay calificaciones
- Glosario HistológicoDocumento4 páginasGlosario HistológicoBeatriz Barco SánchezAún no hay calificaciones
- Especializaciones de La Región Lateral - JONALISDocumento2 páginasEspecializaciones de La Región Lateral - JONALISMichael GonzálezAún no hay calificaciones
- Morfofisiologia Del Tejido CartilaginosoDocumento31 páginasMorfofisiologia Del Tejido CartilaginosobrcoAún no hay calificaciones
- Mapa Mental Tejido ConjuntivoDocumento1 páginaMapa Mental Tejido ConjuntivoHelena SalazarAún no hay calificaciones
- Histología Del Sistema RespiratorioDocumento46 páginasHistología Del Sistema RespiratorioEunice Fabiola ValenzuelaAún no hay calificaciones
- Resumen Tejido Epitelial y ConectivoDocumento5 páginasResumen Tejido Epitelial y ConectivomayraAún no hay calificaciones
- Teoria CelularDocumento16 páginasTeoria Celularkalein100% (2)
- Practica 1 Histología SISTEMA CARDIOVASCULAR y LINFATICO: VASOS SANGUINEOS Y LINFATICOSDocumento11 páginasPractica 1 Histología SISTEMA CARDIOVASCULAR y LINFATICO: VASOS SANGUINEOS Y LINFATICOSPaola Mishell Mamani AyquiAún no hay calificaciones
- Correcto Seminario OJO-OIDODocumento86 páginasCorrecto Seminario OJO-OIDOdanielAún no hay calificaciones
- Tejido EpitelialDocumento13 páginasTejido Epitelialtorresriosjuanantonio52Aún no hay calificaciones
- Tejido EpitelialDocumento7 páginasTejido EpitelialLucy SaucedoAún no hay calificaciones
- Hrubik 2Documento3 páginasHrubik 2Oeste3030Aún no hay calificaciones
- Catalogo Producto FerreteriaDocumento25 páginasCatalogo Producto FerreteriaJuana CortezAún no hay calificaciones
- RESPUESTAS Examen Trimestral Tercer Grado BLOQUE1 2020 2021Documento14 páginasRESPUESTAS Examen Trimestral Tercer Grado BLOQUE1 2020 2021Raúl Jiménez Ocampo100% (1)
- Aplicación Muestreo SistemáticoDocumento17 páginasAplicación Muestreo SistemáticoALEXIS FRANCISCO PINEDA BERNAOLAAún no hay calificaciones
- Dummy 2Documento5 páginasDummy 2Muhammad Al-FatehAún no hay calificaciones
- Los Caballeros de Las GalaxiasDocumento5 páginasLos Caballeros de Las GalaxiasMaria Angélica Morales EcheverriaAún no hay calificaciones
- Esquema Filosofía AnalíticaDocumento2 páginasEsquema Filosofía Analíticamdmma_20100% (2)
- Calidad Del Agua en Los Ecositemas Costeros PDFDocumento407 páginasCalidad Del Agua en Los Ecositemas Costeros PDFBeltran OmarAún no hay calificaciones
- 2009 Señalización Molecular de La Atrofia Por DesusoDocumento11 páginas2009 Señalización Molecular de La Atrofia Por DesusoTote Cifuentes AmigoAún no hay calificaciones
- Estenosis Pilorica A Word ZambranoDocumento18 páginasEstenosis Pilorica A Word Zambranoana luisaAún no hay calificaciones
- Guía 5 Ciclo Iv 2021Documento8 páginasGuía 5 Ciclo Iv 2021Andrea AguilarAún no hay calificaciones
- Manual de Cns en Aps INTERNOSDocumento141 páginasManual de Cns en Aps INTERNOSnicolasrrAún no hay calificaciones
- Fanzine EstoicxsDocumento18 páginasFanzine EstoicxsLeo Vidal100% (1)
- Tomas Austin MillanDocumento12 páginasTomas Austin MillanGladys Galvis Lopez100% (2)
- 2º Quincena 5 AñosDocumento9 páginas2º Quincena 5 AñosPACO MONTOROAún no hay calificaciones
- A.rayder PracticasDocumento4 páginasA.rayder PracticasresAún no hay calificaciones
- Beneficios EmpresarialesDocumento40 páginasBeneficios Empresarialesguille_88Aún no hay calificaciones
- Lista de ParticipantesDocumento2 páginasLista de ParticipanteskrissitoOAún no hay calificaciones
- 01 m2t1 Experto en CitologiaDocumento20 páginas01 m2t1 Experto en CitologiaFelvir RivasAún no hay calificaciones
- Los Chicos de Mi Vida FinalDocumento6 páginasLos Chicos de Mi Vida FinalAnabel Moya LedesmaAún no hay calificaciones
- Guia Modulo 2 - Por Que Se Comete Fraude en Los Estados FinancierosDocumento9 páginasGuia Modulo 2 - Por Que Se Comete Fraude en Los Estados FinancierosCarlos Cuadrado LarreaAún no hay calificaciones
- Lesson 6 Y 7 How To Ask For DirectionsDocumento25 páginasLesson 6 Y 7 How To Ask For Directionsmarynes mundoAún no hay calificaciones
- LibrosDocumento3 páginasLibrosYanira GlezAún no hay calificaciones
- Actividad Matematica #05Documento4 páginasActividad Matematica #05Lessly PicoyAún no hay calificaciones
- 1 Inicial Jassmin Cedeño Planificación Semana #5 Por Experiencias de Aprendizaje # 1 Yo Soy Unico y EspecialDocumento12 páginas1 Inicial Jassmin Cedeño Planificación Semana #5 Por Experiencias de Aprendizaje # 1 Yo Soy Unico y Especialyooomau8Aún no hay calificaciones