También podría gustarte
- Sergio Camargo El Narcotraficante N°82 Alvaro Uribe VelezDocumento284 páginasSergio Camargo El Narcotraficante N°82 Alvaro Uribe VelezJuan Carlos S. Sierra100% (5)
- Maq CorazDocumento1 páginaMaq CorazDaigoro LoveraAún no hay calificaciones
- Seminario Biologia Nº5Documento11 páginasSeminario Biologia Nº5Argenis PeñaAún no hay calificaciones
- Seminario Biologia Nº5Documento11 páginasSeminario Biologia Nº5Argenis PeñaAún no hay calificaciones
- Seminario Biologia Nº8Documento12 páginasSeminario Biologia Nº8Argenis PeñaAún no hay calificaciones
- Seminario Biologia Nº7Documento9 páginasSeminario Biologia Nº7Argenis PeñaAún no hay calificaciones
- Seminario Biologia Nº7Documento9 páginasSeminario Biologia Nº7Argenis PeñaAún no hay calificaciones
- Seminario Biologia Nº9Documento6 páginasSeminario Biologia Nº9Argenis PeñaAún no hay calificaciones
- Seminario Biologia Nº10Documento13 páginasSeminario Biologia Nº10Argenis PeñaAún no hay calificaciones
- Seminario Gentica Nº2Documento10 páginasSeminario Gentica Nº2Argenis PeñaAún no hay calificaciones
- Semi Genetica Nº3Documento8 páginasSemi Genetica Nº3Argenis PeñaAún no hay calificaciones
- Seminario Biologia Nº10Documento13 páginasSeminario Biologia Nº10Argenis PeñaAún no hay calificaciones
- Semi Gen Nº1Documento10 páginasSemi Gen Nº1Argenis PeñaAún no hay calificaciones
- Cartilla BayresPlan 2021Documento21 páginasCartilla BayresPlan 2021Argenis PeñaAún no hay calificaciones
- Aparato Extensor y Biomecanica de Los DedosDocumento9 páginasAparato Extensor y Biomecanica de Los DedosArgenis PeñaAún no hay calificaciones
- Actividad Semanal E6 - 2020Documento16 páginasActividad Semanal E6 - 2020Argenis PeñaAún no hay calificaciones
- Pmo Sanatorios e InternacionDocumento1 páginaPmo Sanatorios e InternacionArgenis PeñaAún no hay calificaciones
- Plan Medico PmoDocumento6 páginasPlan Medico PmoArgenis PeñaAún no hay calificaciones
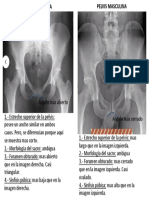
- Diferencias Pelvis Fem y MascDocumento1 páginaDiferencias Pelvis Fem y MascArgenis PeñaAún no hay calificaciones
- BAYRESPLANDocumento7 páginasBAYRESPLANArgenis PeñaAún no hay calificaciones
- Artículo 1º - Modifícase El Anexo I de La Resolución #201Documento6 páginasArtículo 1º - Modifícase El Anexo I de La Resolución #201Argenis PeñaAún no hay calificaciones
- Regiones de La ManoDocumento10 páginasRegiones de La ManoArgenis PeñaAún no hay calificaciones
- Pmo Sanatorios e InternacionDocumento1 páginaPmo Sanatorios e InternacionArgenis PeñaAún no hay calificaciones
- XXDDSDDocumento52 páginasXXDDSDJose Manuel Paz MendozaAún no hay calificaciones
- RELACION M-P y Modelos Medicos 2020Documento11 páginasRELACION M-P y Modelos Medicos 2020Argenis PeñaAún no hay calificaciones
- 4 - Relevamiento de Datos 2020Documento5 páginas4 - Relevamiento de Datos 2020Argenis PeñaAún no hay calificaciones
- Del Positivismo A La ComplejidadDocumento9 páginasDel Positivismo A La ComplejidadArgenis Peña100% (1)
- Trabajo en EquipoDocumento10 páginasTrabajo en EquipoArgenis PeñaAún no hay calificaciones
- 5 - TRANSFER y Contratransf SM 20 (9291)Documento6 páginas5 - TRANSFER y Contratransf SM 20 (9291)Argenis PeñaAún no hay calificaciones
- Tipos de Herencia No MendelianaDocumento22 páginasTipos de Herencia No MendelianaomarcvetAún no hay calificaciones
- BIOTECNOLOGÍA HUMANA UltimoDocumento8 páginasBIOTECNOLOGÍA HUMANA UltimoStevenDanielValverdeAún no hay calificaciones
- Tesis Sara Apaestegui CDocumento68 páginasTesis Sara Apaestegui CFlora JimenezAún no hay calificaciones
- Trabajo Quimica 1102Documento20 páginasTrabajo Quimica 1102Arnold RomeroAún no hay calificaciones
- 10 1007@s12230-020-09801-4 en EsDocumento8 páginas10 1007@s12230-020-09801-4 en EsalvaroAún no hay calificaciones
- Evaluacion BiodiversidadDocumento4 páginasEvaluacion BiodiversidadMarco Antonio Mendoza MolinaAún no hay calificaciones
- PEROXIMOSDocumento7 páginasPEROXIMOSdaniela diazAún no hay calificaciones
- Silabo Biologia 2020-02Documento14 páginasSilabo Biologia 2020-02JRAún no hay calificaciones
- Antonio Banderas Es Medio PlátanoDocumento3 páginasAntonio Banderas Es Medio PlátanoAlejandro Cantarero de SalazarAún no hay calificaciones
- Núcleo 03 Mat 21Documento14 páginasNúcleo 03 Mat 21jhineth yandiAún no hay calificaciones
- 4° Sesion 4Documento4 páginas4° Sesion 4julioAún no hay calificaciones
- Alimentos Transgenicos VELASQUEZDocumento8 páginasAlimentos Transgenicos VELASQUEZYamileth Angela Velásquez Velasquez100% (1)
- Vol3712019 Especial 2019Documento90 páginasVol3712019 Especial 2019GERSON JOEL ORTEGA MORALESAún no hay calificaciones
- A5 AbnerNájera Ensayo MacroevoluciónDocumento4 páginasA5 AbnerNájera Ensayo MacroevoluciónAbner SantesAún no hay calificaciones
- P424204A1033Documento7 páginasP424204A1033jose olayaAún no hay calificaciones
- Taurodontismo TraducidoDocumento8 páginasTaurodontismo TraducidoNadine Madrid ÑaupasAún no hay calificaciones
- Enfoque Biologico y EvolutivoDocumento5 páginasEnfoque Biologico y EvolutivoNatty FerrandoAún no hay calificaciones
- Prueba 4to Año BioDocumento3 páginasPrueba 4to Año BioErnesto PachecoAún no hay calificaciones
- CORRELACIONES CLINICAS CAP 17 - LangmanDocumento4 páginasCORRELACIONES CLINICAS CAP 17 - LangmanLili VanAún no hay calificaciones
- Informe de BiologiaDocumento4 páginasInforme de Biologiatokio den verAún no hay calificaciones
- Anomalías Cromosomicas SexualesDocumento7 páginasAnomalías Cromosomicas SexualesMilenkaGricelAguilarEndaraAún no hay calificaciones
- 3er. Tercer Examen Parcial de Histología Medicina 2017Documento2 páginas3er. Tercer Examen Parcial de Histología Medicina 2017Mar LacrontteAún no hay calificaciones
- Ciencia y Tecnología 10Documento2 páginasCiencia y Tecnología 10Mattias SantosAún no hay calificaciones
- Proteínas de Interés FarmaceuticoDocumento17 páginasProteínas de Interés FarmaceuticoPaulii H ArmenttaAún no hay calificaciones
- Célula - Unidad Orgánica Estructural y FuncionalDocumento1 páginaCélula - Unidad Orgánica Estructural y Funcionalchiara colmanAún no hay calificaciones
- Biotecnología de La Teoría A La Práctica - Recomendaciones para Reportar RECUENTOSDocumento299 páginasBiotecnología de La Teoría A La Práctica - Recomendaciones para Reportar RECUENTOSmiladeraaAún no hay calificaciones
- EXAMEN 1er PARCIAL FISIOLOGIA Y BIOQUIMICADocumento5 páginasEXAMEN 1er PARCIAL FISIOLOGIA Y BIOQUIMICAGus GutiérrezAún no hay calificaciones
- Poke RPGDocumento7 páginasPoke RPGEzequiel FerreyraAún no hay calificaciones
- Actividad 1Documento5 páginasActividad 1mariluz rodriguezAún no hay calificaciones
- EBravo Estructura BiomoleculasDocumento9 páginasEBravo Estructura BiomoleculasKAROL GUTIERREZ MADRIGALAún no hay calificaciones