También podría gustarte
- FisiologiaDocumento9 páginasFisiologiaRoxany ValeriaAún no hay calificaciones
- FisiologiaDocumento8 páginasFisiologiaRoxany ValeriaAún no hay calificaciones
- Epidemiología de La Peste NegraDocumento3 páginasEpidemiología de La Peste NegraRoxany ValeriaAún no hay calificaciones
- Elaboración de Un PerfumeDocumento28 páginasElaboración de Un PerfumeRoxany ValeriaAún no hay calificaciones
- Pratica Gasto CardiacoDocumento5 páginasPratica Gasto CardiacoIvonne CecibeAún no hay calificaciones
- Precarga y Post CargaDocumento3 páginasPrecarga y Post CargaMaria SalazarAún no hay calificaciones
- Ciclo Cardiaco SemiDocumento88 páginasCiclo Cardiaco SemiKaori Coral SalvadorAún no hay calificaciones
- Fisiologia Cardiovascular Pediatrica PDFDocumento42 páginasFisiologia Cardiovascular Pediatrica PDFManuel Azabache100% (9)
- Monitoreo HemodinámicoDocumento5 páginasMonitoreo HemodinámicoKarimAún no hay calificaciones
- FurosemidaDocumento7 páginasFurosemidaCarlos Alberto Angel GutiérrezAún no hay calificaciones
- INTERACCIÓN CORAZÓN PULMÓN-editadoDocumento8 páginasINTERACCIÓN CORAZÓN PULMÓN-editadoJulian PaguanquizaAún no hay calificaciones
- Sistema CardiovascularDocumento50 páginasSistema CardiovascularLuz GarizadoAún no hay calificaciones
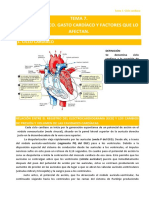
- Tema 7. Ciclo Cardíaco. Gasto Cardíaco y Factores Que LoDocumento17 páginasTema 7. Ciclo Cardíaco. Gasto Cardíaco y Factores Que LoRosa SotoAún no hay calificaciones
- Informe Laboratorio..Documento9 páginasInforme Laboratorio..Valentina MendezAún no hay calificaciones
- CardiologiaDocumento55 páginasCardiologiadianaAún no hay calificaciones
- Fisiopatologia Cardiovascular CompendioDocumento47 páginasFisiopatologia Cardiovascular CompendioAlexandra AraujoAún no hay calificaciones
- Gasto CardiacoDocumento7 páginasGasto CardiacoAnyey AcuñaAún no hay calificaciones
- Sitema CardiovascularDocumento13 páginasSitema CardiovascularRo AvilaAún no hay calificaciones
- Universidad de CaraboboDocumento4 páginasUniversidad de CaraboboDon AlbertoAún no hay calificaciones
- MODULO 12 ANATOMIA - Voxel en Cardiologia. Manual de Patologia Por Imagen. - ESPAÑOLDocumento183 páginasMODULO 12 ANATOMIA - Voxel en Cardiologia. Manual de Patologia Por Imagen. - ESPAÑOLEliana Solange FouzAún no hay calificaciones
- Copia de Guía N 14 - Ciclo Cardíaco y Regulación Del Volumen Minuto. 2021. SIN RESOLVERDocumento29 páginasCopia de Guía N 14 - Ciclo Cardíaco y Regulación Del Volumen Minuto. 2021. SIN RESOLVERMalena EcheverríaAún no hay calificaciones
- Tresguerres Capitulo 38 - FisiologiaDocumento37 páginasTresguerres Capitulo 38 - FisiologiaEric KirAún no hay calificaciones
- Corazón2 2 1Documento47 páginasCorazón2 2 1Gabyta BarreraAún no hay calificaciones
- Insuficiencia CardiacaDocumento10 páginasInsuficiencia CardiacaPaul CabreraAún no hay calificaciones
- La Bomba Cardíaca Ciclo Cardíaco Regulación P. ArterialDocumento43 páginasLa Bomba Cardíaca Ciclo Cardíaco Regulación P. ArterialRazec694Aún no hay calificaciones
- Charla Fisiologia-1Documento3 páginasCharla Fisiologia-1Sara MoranAún no hay calificaciones
- Cuestionario Cardio Copia A Los AlumnosDocumento4 páginasCuestionario Cardio Copia A Los AlumnosAnonymous i6XY1SG0% (1)
- Glucósidos CardiotónicosDocumento52 páginasGlucósidos CardiotónicosRobin MalpartidaAún no hay calificaciones
- Musculo LisoDocumento6 páginasMusculo LisoMaurimar LarezAún no hay calificaciones
- Fisiopatología Del Sistema Cardiovascular Y Respiratorio.Documento54 páginasFisiopatología Del Sistema Cardiovascular Y Respiratorio.maria isabel briceñoAún no hay calificaciones
- Potenciales de Marcapasos y MiocardiocitosDocumento14 páginasPotenciales de Marcapasos y MiocardiocitosJcaminoAún no hay calificaciones
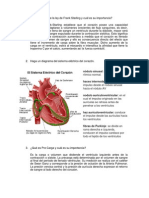
- En Qué Consiste La Ley de Frank Starling y Cuál Es Su ImportanciaDocumento2 páginasEn Qué Consiste La Ley de Frank Starling y Cuál Es Su ImportanciaKatha Ramos79% (28)
- POTENCIA DE MARCAPASOS-s10Documento14 páginasPOTENCIA DE MARCAPASOS-s10Mariela Anton Chunga100% (1)
- Frank StarlingDocumento3 páginasFrank StarlingAnita ApazaAún no hay calificaciones
- Homo antecessor: El nacimiento de una especieDe EverandHomo antecessor: El nacimiento de una especieCalificación: 5 de 5 estrellas5/5 (1)
- 50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaDe Everand50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaCalificación: 4 de 5 estrellas4/5 (37)
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)De EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Calificación: 5 de 5 estrellas5/5 (9)
- El psoas: Músculo vital (Color)De EverandEl psoas: Músculo vital (Color)Calificación: 3.5 de 5 estrellas3.5/5 (9)
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- Sesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesDe EverandSesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesCalificación: 4.5 de 5 estrellas4.5/5 (13)
- Cerebro y silencio: Las claves de la creatividad y la serenidadDe EverandCerebro y silencio: Las claves de la creatividad y la serenidadCalificación: 5 de 5 estrellas5/5 (2)
- Liberación del trauma: Perdón y temblor es el caminoDe EverandLiberación del trauma: Perdón y temblor es el caminoCalificación: 4 de 5 estrellas4/5 (4)
- La comida y el inconsciente: Psicoanálisis y trastornos alimentariosDe EverandLa comida y el inconsciente: Psicoanálisis y trastornos alimentariosAún no hay calificaciones
- Cerebros rotos: Pacientes asombrosos que me enseñaron a vivirDe EverandCerebros rotos: Pacientes asombrosos que me enseñaron a vivirCalificación: 5 de 5 estrellas5/5 (3)
- Conciencia del tiempo: Por qué pensar como geólogos puede ayudarnos a salvar el planetaDe EverandConciencia del tiempo: Por qué pensar como geólogos puede ayudarnos a salvar el planetaCalificación: 4.5 de 5 estrellas4.5/5 (2)
- 200 tareas en terapia breve: 2ª ediciónDe Everand200 tareas en terapia breve: 2ª ediciónCalificación: 4.5 de 5 estrellas4.5/5 (33)
- Historia del cuerpo humano: Evolución, salud y enfermedadDe EverandHistoria del cuerpo humano: Evolución, salud y enfermedadAún no hay calificaciones
- Cómo estimular el cerebro del niño: 100 ejercicios para potenciar la concentración, la memoria y otras funciones ejecutivasDe EverandCómo estimular el cerebro del niño: 100 ejercicios para potenciar la concentración, la memoria y otras funciones ejecutivasCalificación: 4.5 de 5 estrellas4.5/5 (12)