También podría gustarte
- Qué es la Epilepsia. Causas, Síntomas, Diagnóstico y Tratamiento: TRASTORNOS DEL SUEÑO, #2De EverandQué es la Epilepsia. Causas, Síntomas, Diagnóstico y Tratamiento: TRASTORNOS DEL SUEÑO, #2Aún no hay calificaciones
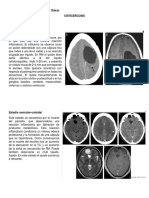
- EVC Isquémico FisiopatologiaDocumento58 páginasEVC Isquémico FisiopatologiaSalvador FigueroaAún no hay calificaciones
- Enfermedad Articular DegenerativaDocumento16 páginasEnfermedad Articular DegenerativaAnahi MartinezAún no hay calificaciones
- Absorción con filtro BK como terapia eficaz en el riñón del mieloma múltipleDe EverandAbsorción con filtro BK como terapia eficaz en el riñón del mieloma múltipleAún no hay calificaciones
- Enfermo GeriátricoDocumento9 páginasEnfermo GeriátricoROSI100% (1)
- Sindromes GeriátricosDocumento2 páginasSindromes GeriátricosAleja TorresAún no hay calificaciones
- Tratamiento de Lesiones Intraepiteliales Escamosas Cervico Vaginales.Documento5 páginasTratamiento de Lesiones Intraepiteliales Escamosas Cervico Vaginales.OliverAún no hay calificaciones
- Presentacion de EpocDocumento22 páginasPresentacion de EpocNidia Isabel BunburyAún no hay calificaciones
- Anatomía Del Oido ResumenDocumento3 páginasAnatomía Del Oido ResumenMiriam100% (1)
- Aparato yuxtaglomerular y sus célulasDocumento14 páginasAparato yuxtaglomerular y sus célulasMedtercersemestre Tominaga NakamotoAún no hay calificaciones
- Protocolo de RCP en Mujeres EmbarazadasDocumento3 páginasProtocolo de RCP en Mujeres EmbarazadasGema Hermida100% (2)
- Traumatismo Craneo EncefalicoDocumento100 páginasTraumatismo Craneo EncefalicoVictor Daniel Pelagio Quintana100% (1)
- Manual cuidador adulto mayor dependiente síndromesDocumento29 páginasManual cuidador adulto mayor dependiente síndromesAnabel IpialesAún no hay calificaciones
- Artritis Juvenil Idiopatica Rehabilitacion Y ReadaptacionDocumento15 páginasArtritis Juvenil Idiopatica Rehabilitacion Y ReadaptacionMabel BonifacioAún no hay calificaciones
- NeurocisticercosisDocumento2 páginasNeurocisticercosisMaira Vargas OdicioAún no hay calificaciones
- 15 Rehabilitación Del Adulto MayorDocumento7 páginas15 Rehabilitación Del Adulto MayorXyxnxmiAún no hay calificaciones
- Mieloma Multiple Traumatologia 5..Documento23 páginasMieloma Multiple Traumatologia 5..Chai AlvarezAún no hay calificaciones
- Síndromes GeriátricosDocumento13 páginasSíndromes GeriátricosDiane Troncoso AlegriaAún no hay calificaciones
- Presentacion IctusDocumento30 páginasPresentacion IctusGuaci 153Aún no hay calificaciones
- Urgencias Traumatologicas y OrtopedicasDocumento9 páginasUrgencias Traumatologicas y OrtopedicasFrancy Luc Martinez RojasAún no hay calificaciones
- Lesiones Vasculares de Miembros Inferiores PressentacioDocumento56 páginasLesiones Vasculares de Miembros Inferiores PressentacioJude AccouAún no hay calificaciones
- Injertos Óseos y Sus SustitutosDocumento9 páginasInjertos Óseos y Sus SustitutosErick AlexanderAún no hay calificaciones
- Guia Hematoma SubduralDocumento4 páginasGuia Hematoma SubduralÁngel M. JeyAún no hay calificaciones
- Tumor de WarthinDocumento6 páginasTumor de WarthinAndyRamosLopezAún no hay calificaciones
- Laringitis y sus causasDocumento22 páginasLaringitis y sus causasElizabeth Mendoza EspinosaAún no hay calificaciones
- Torax Inestable y HemomediastinoDocumento19 páginasTorax Inestable y Hemomediastinobrenda marquezAún no hay calificaciones
- Cefalea y Neuralgias 1.1Documento54 páginasCefalea y Neuralgias 1.1Ana Guadalupe Montiel juarezAún no hay calificaciones
- Fiebre Reumática PresentaciónDocumento37 páginasFiebre Reumática Presentacióningenierojesus2014Aún no hay calificaciones
- Artropatia DegererativaDocumento21 páginasArtropatia DegererativaRosarioAún no hay calificaciones
- APENDICECTOMIADocumento8 páginasAPENDICECTOMIAAngie BautistaAún no hay calificaciones
- Hematologia Place FinalDocumento36 páginasHematologia Place Finaldiego de jesus lara garciaAún no hay calificaciones
- Fibrilacion AuricularDocumento141 páginasFibrilacion AuricularMiile OrozcoAún no hay calificaciones
- Generalidades de las Enfermedades ReumáticasDocumento26 páginasGeneralidades de las Enfermedades ReumáticasMiguel CalderonAún no hay calificaciones
- Trauma torácico: evaluación y manejo inicial de lesiones que comprometen la vidaDocumento6 páginasTrauma torácico: evaluación y manejo inicial de lesiones que comprometen la vidaNadia BarrónAún no hay calificaciones
- Enfermedad de HuntingtonDocumento4 páginasEnfermedad de HuntingtonDanae TatajeAún no hay calificaciones
- Cambios en La Piel Del Geronte PDFDocumento9 páginasCambios en La Piel Del Geronte PDFIvanovich NickAún no hay calificaciones
- Enfermedad Carotídea PDFDocumento23 páginasEnfermedad Carotídea PDFMarisolCamposAún no hay calificaciones
- Tumores Oculares Más FrecuentesDocumento68 páginasTumores Oculares Más FrecuentesJesus Efren Villa GarciaAún no hay calificaciones
- Cardiopatía IsquémicaDocumento44 páginasCardiopatía IsquémicacelesteAún no hay calificaciones
- Vías Aéreas Superiores e Inferiores, Circulación Sanguínea A Nivel PulmonarDocumento75 páginasVías Aéreas Superiores e Inferiores, Circulación Sanguínea A Nivel PulmonarMishypaiAún no hay calificaciones
- Detecta síndromes geriátricos con SPICESDocumento20 páginasDetecta síndromes geriátricos con SPICESbeibeibei100% (1)
- VitiligoDocumento19 páginasVitiligoNadia PDAún no hay calificaciones
- Teorías del envejecimiento: gerodinámica, desvinculación y medio socialDocumento7 páginasTeorías del envejecimiento: gerodinámica, desvinculación y medio socialPerla GonzalezAún no hay calificaciones
- Fragilidad y CaídasDocumento51 páginasFragilidad y CaídasSteffany LopezAún no hay calificaciones
- Encefalopatia HepaticaDocumento4 páginasEncefalopatia HepaticaDavid PazAún no hay calificaciones
- Pae Hemodialisis.Documento24 páginasPae Hemodialisis.Jesus GarciaAún no hay calificaciones
- Protocolo de HipokalemiaDocumento14 páginasProtocolo de HipokalemiaKATHERINE LEONAún no hay calificaciones
- Trauma RaquimedularDocumento10 páginasTrauma RaquimedularAna Maria VegaAún no hay calificaciones
- Proceso de Envejecimiento, Teorias de Envejecimiento y Adulto Mayor Sano PDFDocumento43 páginasProceso de Envejecimiento, Teorias de Envejecimiento y Adulto Mayor Sano PDFandres sotoAún no hay calificaciones
- ParotiditisDocumento8 páginasParotiditisLalo Gonzalez LubckeAún no hay calificaciones
- Pancreatitis Caso ClínicoDocumento3 páginasPancreatitis Caso ClínicoAngel Choque VelásquezAún no hay calificaciones
- Sindrome de LI FRAUMENIDocumento16 páginasSindrome de LI FRAUMENIErika itzel LimaAún no hay calificaciones
- Esclerosis MultipleDocumento22 páginasEsclerosis MultipleKarolina Quintana Hermosilla100% (1)
- HEMONEUMOTORAXDocumento3 páginasHEMONEUMOTORAXDahiana L. NúñezAún no hay calificaciones
- Libro Comppleto Enfs. Reumaticas y EmbarazoDocumento300 páginasLibro Comppleto Enfs. Reumaticas y Embarazomanuel ignacio segura martinezAún no hay calificaciones
- Trastornos de Salud Adulto MayorDocumento32 páginasTrastornos de Salud Adulto MayorGenesisAún no hay calificaciones
- Envejecimiento FisiológicoDocumento24 páginasEnvejecimiento FisiológicoAivree DreowoneAún no hay calificaciones
- Injertos de PielDocumento8 páginasInjertos de PielPatty MoretaAún no hay calificaciones
- UlcerasDocumento5 páginasUlcerasJuan David AlvarezAún no hay calificaciones
- Cuestionario - Semana 11Documento3 páginasCuestionario - Semana 11Yerlin Huamán BautistaAún no hay calificaciones
- SEMIN - Medicina HipocráticaDocumento25 páginasSEMIN - Medicina HipocráticaYerlin Huamán BautistaAún no hay calificaciones
- BiomoléculasDocumento8 páginasBiomoléculasYerlin Huamán BautistaAún no hay calificaciones
- Costanzo - Fisiología - Preguntas y Respuestas (Alternativas)Documento110 páginasCostanzo - Fisiología - Preguntas y Respuestas (Alternativas)Luz M. Martín100% (1)
- Escala de TannerDocumento2 páginasEscala de TannerLuis Angel AmezcuaAún no hay calificaciones
- Metodos Anticonceptivos FinalDocumento29 páginasMetodos Anticonceptivos FinalYerlin Huamán BautistaAún no hay calificaciones
- Desempeño 3Documento17 páginasDesempeño 3Yerlin Huamán BautistaAún no hay calificaciones
- Lectura 3aDocumento16 páginasLectura 3aYerlin Huamán BautistaAún no hay calificaciones
- Quimica-Propiedades de La Materia: Densidad y ViscosidadDocumento11 páginasQuimica-Propiedades de La Materia: Densidad y ViscosidadYerlin Huamán BautistaAún no hay calificaciones
- La Historia Clínica. Marco LegalDocumento6 páginasLa Historia Clínica. Marco LegalSamuel Flores CalderonAún no hay calificaciones
- Cuadros Reflexion BCEPDocumento11 páginasCuadros Reflexion BCEPnapsitaxAún no hay calificaciones
- Couma Macrocarpa OsinforDocumento2 páginasCouma Macrocarpa OsinforWilliam Reynaldo Ñahuero JulcarimaAún no hay calificaciones
- S1-Tarea 1.1-Cadena de SuministrosDocumento7 páginasS1-Tarea 1.1-Cadena de SuministrosKarol PinedaAún no hay calificaciones
- Acertijos LogicosDocumento3 páginasAcertijos LogicosferdinandcruzAún no hay calificaciones
- Manual Curiso GisDocumento98 páginasManual Curiso GisRichard2023Aún no hay calificaciones
- Instrumentos para Medición Angular y LinealDocumento27 páginasInstrumentos para Medición Angular y LinealEl aleczis50% (2)
- Exp3 Secundaria 3y4 Seguimosaprendiendo EducacionparaeltrabajoAct03IdeamoslasolucionDocumento4 páginasExp3 Secundaria 3y4 Seguimosaprendiendo EducacionparaeltrabajoAct03Ideamoslasolucionjorge castañedaAún no hay calificaciones
- Evidencia 3 Informe Resultados FinancierosDocumento13 páginasEvidencia 3 Informe Resultados FinancierosSTEVEN AMORTEGUI100% (1)
- Sumativa 4 - Solemne 1.0 ..Documento3 páginasSumativa 4 - Solemne 1.0 ..nelsonAún no hay calificaciones
- Tesis de Arrascue Delgado y Segura CardozoDocumento150 páginasTesis de Arrascue Delgado y Segura CardozoLucia DazaAún no hay calificaciones
- Innovaciones Modernas en La Construccion de Pavimentos FlexiblesDocumento12 páginasInnovaciones Modernas en La Construccion de Pavimentos FlexiblesEduardoChavezAún no hay calificaciones
- Practica de AlgoritmosDocumento1 páginaPractica de AlgoritmosAndrey SolanoAún no hay calificaciones
- Memorias Fundición AceroDocumento10 páginasMemorias Fundición AceroRICARDO ANDRES CASTRO ROJASAún no hay calificaciones
- Cuestionario Unidad 2Documento12 páginasCuestionario Unidad 2hilbert_326100% (1)
- Desarrollo Urbano y Ordenamiento Territorial La LibertadDocumento38 páginasDesarrollo Urbano y Ordenamiento Territorial La LibertadSheyla Gutierrez BarrientosAún no hay calificaciones
- GinsburgDocumento8 páginasGinsburgVittoria CorleoneAún no hay calificaciones
- UCV-2022-2 - MIC-C28 - Grupo 1 - 2022-11-4Documento19 páginasUCV-2022-2 - MIC-C28 - Grupo 1 - 2022-11-4KIARA FERNANDA ROMANI JARAAún no hay calificaciones
- EJERCICIOS PROPUESTOS Segunda UnidadDocumento2 páginasEJERCICIOS PROPUESTOS Segunda UnidadRocio ObregonAún no hay calificaciones
- Norma Ecuatoriana de Determinacion de ColorDocumento14 páginasNorma Ecuatoriana de Determinacion de ColorPaam InstrumentalAún no hay calificaciones
- Cultivo de la okra: descripción botánica, requerimientos y manejoDocumento24 páginasCultivo de la okra: descripción botánica, requerimientos y manejoernesto34346Aún no hay calificaciones
- Sesion de Inicial 5 AñosDocumento6 páginasSesion de Inicial 5 AñosElver Farfan InafnteAún no hay calificaciones
- Reglamento de Correspondencia y de ArchivoDocumento28 páginasReglamento de Correspondencia y de ArchivoFonseca MileAún no hay calificaciones
- Análisis de los héroes clásico, medieval y renacentistaDocumento4 páginasAnálisis de los héroes clásico, medieval y renacentistajuan1212Aún no hay calificaciones
- 12.1 - Anatomía Clínica de Los PárpadosDocumento5 páginas12.1 - Anatomía Clínica de Los PárpadosSary Rivadeneira100% (1)
- Cronograma Prácticos - LUNESDocumento5 páginasCronograma Prácticos - LUNESjtvaldes100% (1)
- Biografia de Carlos NoriegaDocumento5 páginasBiografia de Carlos NoriegaLuis Apaza100% (2)
- Unidad 1. Programacion LinealDocumento16 páginasUnidad 1. Programacion LinealAlessandro BarredaAún no hay calificaciones
- Guía de aprendizaje de seguridad industrialDocumento48 páginasGuía de aprendizaje de seguridad industrialpedro manuel antequera castroAún no hay calificaciones