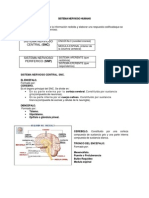
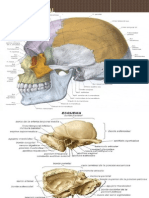
También podría gustarte
- Potencial de Reposo y EquilibrioDocumento5 páginasPotencial de Reposo y EquilibrioDan PariascaAún no hay calificaciones
- Cilios y FlagelosDocumento6 páginasCilios y FlagelosXavi Zambrano GIAún no hay calificaciones
- Sinapsis. Quimica y Electrica ppt4Documento8 páginasSinapsis. Quimica y Electrica ppt4Camilo San Martin CarrascoAún no hay calificaciones
- Sistema Muscular Del Cuerpo Humano y Sus PartesDocumento5 páginasSistema Muscular Del Cuerpo Humano y Sus PartesLaura CastilloAún no hay calificaciones
- URETRADocumento10 páginasURETRAcamachitaAún no hay calificaciones
- Fisiología NeuromuscularDocumento5 páginasFisiología NeuromuscularOscar Placencia AgredaAún no hay calificaciones
- Modelo Electrico de La Menbrana CelularDocumento8 páginasModelo Electrico de La Menbrana CelularAlex GarciaAún no hay calificaciones
- Pares Craneales y Su FuncionDocumento25 páginasPares Craneales y Su FuncionRobinsonGiraldoLópez100% (1)
- Capitulo Señalizacion Celular-ReceptoresDocumento60 páginasCapitulo Señalizacion Celular-ReceptoresEdwar Leyva Baltazar0% (1)
- Intro Historia de Los AnestésicosDocumento12 páginasIntro Historia de Los AnestésicosGonzalo Paredes GonzalesAún no hay calificaciones
- Cuestionario de Tejido NerviosoDocumento2 páginasCuestionario de Tejido NerviosoAna Karen Aguilar RicoAún no hay calificaciones
- 03 - Alteraciones de La ConcienciaDocumento34 páginas03 - Alteraciones de La ConcienciaAle ZambranoAún no hay calificaciones
- Region CigomaticaDocumento35 páginasRegion CigomaticaAJ_evaAún no hay calificaciones
- Generacion y Conduccion de Potenciales de AcciónDocumento8 páginasGeneracion y Conduccion de Potenciales de AcciónIván Andrés Escobar Figueroa0% (1)
- Sistematización Anatómica de La Médula EspinalDocumento3 páginasSistematización Anatómica de La Médula EspinalVictorFloresAnchanteAún no hay calificaciones
- Histologia Del CorazonDocumento16 páginasHistologia Del Corazonlaura katalina mejia rodriguezAún no hay calificaciones
- Cap 47 Sensibilidades Somáticas - I. Organización General, Las Sensaciones Táctil y Posicional PDFDocumento8 páginasCap 47 Sensibilidades Somáticas - I. Organización General, Las Sensaciones Táctil y Posicional PDFrodrigo4590100% (1)
- BiofluidosDocumento6 páginasBiofluidosAndresAún no hay calificaciones
- Potencial de Acción - FisiologíaDocumento6 páginasPotencial de Acción - FisiologíaDara DosAún no hay calificaciones
- Canales IónicosDocumento12 páginasCanales IónicosJulinho M LeOnAún no hay calificaciones
- 20 LaringitisDocumento9 páginas20 LaringitisLilo VotesAún no hay calificaciones
- HipotalamoDocumento4 páginasHipotalamoIrvin Mtz AristaAún no hay calificaciones
- Tejido Epitelial Usmp HistologiaDocumento63 páginasTejido Epitelial Usmp Histologiavibescash100% (1)
- Enfoques PsicologicosDocumento19 páginasEnfoques PsicologicosholasdasdsAún no hay calificaciones
- SubtalamoDocumento50 páginasSubtalamoMariand F Morales100% (2)
- Importancia Del Sistema de PurkinjeDocumento1 páginaImportancia Del Sistema de PurkinjeCeovi VillanAún no hay calificaciones
- Lipotimia Shock Anafilactico y SincopeDocumento25 páginasLipotimia Shock Anafilactico y SincopeDarwin Mamani ReyesAún no hay calificaciones
- Region Fosa Cigomatica ExpoDocumento14 páginasRegion Fosa Cigomatica ExpoElvis Ponce ColquiAún no hay calificaciones
- Glandula Submandibular.Documento7 páginasGlandula Submandibular.Marcos Erli Banda CubasAún no hay calificaciones
- Nuevo Documento de Microsoft Office WordDocumento5 páginasNuevo Documento de Microsoft Office WordRube Garduño RoaAún no hay calificaciones
- Tarea Sinapsis y Union NeuromuscularDocumento4 páginasTarea Sinapsis y Union NeuromuscularOrlando ComtrerazAún no hay calificaciones
- Señalizacion Celular y Sistema Endocrino PDFDocumento27 páginasSeñalizacion Celular y Sistema Endocrino PDFFiorella Alzamora VignaAún no hay calificaciones
- Anexo de Irrigación y Inervación de ATMDocumento1 páginaAnexo de Irrigación y Inervación de ATMGonzalo San MartinAún no hay calificaciones
- Morfofisiología de La NeuronaDocumento3 páginasMorfofisiología de La Neuronami bella mas bellaAún no hay calificaciones
- Informe Re GolgiDocumento7 páginasInforme Re GolgienriqueAún no hay calificaciones
- Potencial de Accion y MembranaDocumento2 páginasPotencial de Accion y MembranaEmily Ortegon100% (1)
- Artículo Científico de MicrosDocumento13 páginasArtículo Científico de MicrosDANIEL ANGEL TORRES ESTUDIANTE ACTIVOAún no hay calificaciones
- Cuestionario Tejido NerviosoDocumento6 páginasCuestionario Tejido NerviosoAna BazzanoAún no hay calificaciones
- Termodinámica Aplicada Al HombreDocumento11 páginasTermodinámica Aplicada Al HombreLuis Enrique ArrietaInfanteAún no hay calificaciones
- Vía TrigeminalDocumento11 páginasVía TrigeminalDANIEL ALEJANDRO88% (8)
- 3.ppse y PpsiDocumento63 páginas3.ppse y PpsiJOhn JacoboAún no hay calificaciones
- Resumen de MorfofuncionDocumento8 páginasResumen de MorfofuncionPoly Cab100% (1)
- Historia Natural de La EnfermedadDocumento5 páginasHistoria Natural de La EnfermedadShaid Castañeda YbarrolaAún no hay calificaciones
- Pares Craneales o Nervios CranealesDocumento10 páginasPares Craneales o Nervios CranealesAntonio Arroyo CornejoAún no hay calificaciones
- Sentidos EspecialesDocumento8 páginasSentidos EspecialesIon Frias0% (1)
- Anatomia y Fisiologia Del Sistema NerviosoDocumento6 páginasAnatomia y Fisiologia Del Sistema NerviosoCristian AcostaAún no hay calificaciones
- Informe (Cavidad Nasal) - Exposiciòn (Oìdo Medio)Documento21 páginasInforme (Cavidad Nasal) - Exposiciòn (Oìdo Medio)Mary Monja DiazAún no hay calificaciones
- Nervio Vestibulococlear, AnatomíaDocumento3 páginasNervio Vestibulococlear, AnatomíaMartín MeléndezAún no hay calificaciones
- Caso Clinico Clase VDocumento18 páginasCaso Clinico Clase VGisselle CaceresAún no hay calificaciones
- Sistema Nervioso TripticoDocumento2 páginasSistema Nervioso TripticoMariedjalex MarieAún no hay calificaciones
- Neuro Transm I SoresDocumento5 páginasNeuro Transm I Soresdamaris pradoAún no hay calificaciones
- Realizar Un Mapa ConceptualDocumento6 páginasRealizar Un Mapa ConceptualjuanAún no hay calificaciones
- Musculo LisoDocumento9 páginasMusculo LisoMartinVelazcoAún no hay calificaciones
- Cap 48 GuytonDocumento4 páginasCap 48 GuytonCinthia LemusAún no hay calificaciones
- Areas de Brodman de La Corteza HumanaDocumento9 páginasAreas de Brodman de La Corteza HumanaJorge Manuel Pisco ZavaletaAún no hay calificaciones
- Fosa TemporalDocumento18 páginasFosa TemporalJosurer NrtAún no hay calificaciones
- Neuro CompletoDocumento162 páginasNeuro CompletoBrenda PusdáAún no hay calificaciones
- Potencial de Membrana en Reposo de Células ExcitablesDocumento13 páginasPotencial de Membrana en Reposo de Células ExcitablesDaniel SantanaAún no hay calificaciones
- Clase Q.F. Alejandra Del Rio ISP Registro Productos Farmaceuticos Biologicos 2021Documento58 páginasClase Q.F. Alejandra Del Rio ISP Registro Productos Farmaceuticos Biologicos 2021ScarlethMuñozAún no hay calificaciones
- Clase 4 Intervalos de ConfianzaDocumento56 páginasClase 4 Intervalos de ConfianzaScarlethMuñozAún no hay calificaciones
- Clase 2Documento48 páginasClase 2ScarlethMuñozAún no hay calificaciones
- Resumen de Técnicas de LaboratorioDocumento20 páginasResumen de Técnicas de LaboratorioScarlethMuñozAún no hay calificaciones
- Infección Urinaria y Gestación: Protocolos SEGODocumento6 páginasInfección Urinaria y Gestación: Protocolos SEGOManuel VasquezAún no hay calificaciones
- Bases de Las Relaciones ÍntimasDocumento5 páginasBases de Las Relaciones ÍntimasFlavii Estefanii ST50% (2)
- Informe de BiologiaDocumento6 páginasInforme de BiologiaJorge Luis Manzaba MoralesAún no hay calificaciones
- Encimas CardiacasDocumento43 páginasEncimas CardiacasJorge MTz OcampoAún no hay calificaciones
- EMBALSAMAMIENTODocumento4 páginasEMBALSAMAMIENTOAndrea D'OnofrioAún no hay calificaciones
- Diaz Navarro, WendyDocumento41 páginasDiaz Navarro, WendyRodríguez Rafael EdelmiroAún no hay calificaciones
- FitopatologiaDocumento33 páginasFitopatologiajosetorrealba84100% (1)
- ApotemnofiliaDocumento2 páginasApotemnofiliaMaria Jose MorenoAún no hay calificaciones
- Sedentarismo en Adultos MayoresDocumento4 páginasSedentarismo en Adultos MayoresalisonAún no hay calificaciones
- Embriologia General.Documento9 páginasEmbriologia General.Candela Carrion BustosAún no hay calificaciones
- Drenaje Linfatico Manual (69 Pag)Documento70 páginasDrenaje Linfatico Manual (69 Pag)Andrea Marcela Sáez D'ArcangeliAún no hay calificaciones
- Tratamiento para DermatitisDocumento4 páginasTratamiento para DermatitisDickson Dib GalebAún no hay calificaciones
- Consulta PreconcepcionalDocumento7 páginasConsulta PreconcepcionalLaEneydaAún no hay calificaciones
- Homicidio en CelebritiesDocumento7 páginasHomicidio en CelebritiesjuanAún no hay calificaciones
- Diagnostico de DepresionDocumento8 páginasDiagnostico de DepresionAnonymous yRO7LCAún no hay calificaciones
- Segundos Mensajeros Bien EstablecidosDocumento2 páginasSegundos Mensajeros Bien EstablecidosWesttler Emanuel AlonzoAún no hay calificaciones
- Antioxidantes Reguladores de PH DiapositivasDocumento24 páginasAntioxidantes Reguladores de PH DiapositivasJulio Cesar Torres PereyraAún no hay calificaciones
- Nuevo Sistema de Avisos de Accidentes de Trabajo SIAATDocumento5 páginasNuevo Sistema de Avisos de Accidentes de Trabajo SIAATLucy UgarteAún no hay calificaciones
- Folleto ObesidadDocumento2 páginasFolleto ObesidadHospital de Alta Complejidad "Pte. juan DOmingo Perón"Aún no hay calificaciones
- Plantas Medicinales Hierba Santa RudaDocumento3 páginasPlantas Medicinales Hierba Santa Rudakarlosdan92Aún no hay calificaciones
- Mammuth UsDocumento12 páginasMammuth UsRicardo CastroAún no hay calificaciones
- Bioquimica ResumenDocumento13 páginasBioquimica ResumenEstefanykeiko ReyesAquinoAún no hay calificaciones
- Clase 10 Sistema Regulador NerviosoDocumento27 páginasClase 10 Sistema Regulador NerviosogersonAún no hay calificaciones
- BGNFDocumento15 páginasBGNFCristhian A. E.R.Aún no hay calificaciones
- Principios de Neuroanatomia PDFDocumento2 páginasPrincipios de Neuroanatomia PDFOscar TorresAún no hay calificaciones
- Consecuencias de Una Mala AlimentacionDocumento25 páginasConsecuencias de Una Mala AlimentacionEmanuel Díez100% (1)
- EPI INFO Tema de InvestigacionDocumento11 páginasEPI INFO Tema de InvestigacionMARCEOAún no hay calificaciones
- Manual Toma MuestraDocumento48 páginasManual Toma MuestraJalea HumanaAún no hay calificaciones
- 42-Texto Del Artículo-126-3-10-20200804Documento59 páginas42-Texto Del Artículo-126-3-10-20200804ChaveliAún no hay calificaciones
- Grupos SanguineosDocumento35 páginasGrupos SanguineosClemencia Alvarez VillaAún no hay calificaciones