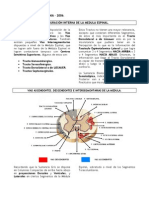
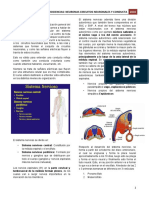
También podría gustarte
- Teoría Del Rol FamiliaDocumento23 páginasTeoría Del Rol FamiliaLlacsa yessica100% (1)
- Depósitos IOCGDocumento41 páginasDepósitos IOCGD'Marlon MnAún no hay calificaciones
- Seminario Biopotenciales FisiologiaDocumento9 páginasSeminario Biopotenciales Fisiologiayamirjossue100% (9)
- Aparato CardiovascularDocumento11 páginasAparato CardiovascularMel RoniAún no hay calificaciones
- Fibra Óptica Sesiones 1-6Documento124 páginasFibra Óptica Sesiones 1-6dgonzalezhAún no hay calificaciones
- Teoria Aprendizaje Significativo 29075Documento85 páginasTeoria Aprendizaje Significativo 29075Manuel Portocarrero VargasAún no hay calificaciones
- Capitulo 4 Introduccion A Fibras OpticasDocumento104 páginasCapitulo 4 Introduccion A Fibras OpticasRebeca OrtizAún no hay calificaciones
- Composición Del Cuerpo HumanoDocumento3 páginasComposición Del Cuerpo HumanoJohnny MosquedaAún no hay calificaciones
- Cable de Fibra ÓpticaDocumento4 páginasCable de Fibra ÓpticaAlejandro SarmientoAún no hay calificaciones
- Manual de Uso de Radio MobileDocumento18 páginasManual de Uso de Radio MobileÁngel Castro SánchezAún no hay calificaciones
- Tarea Sinapsis y Union NeuromuscularDocumento4 páginasTarea Sinapsis y Union NeuromuscularOrlando ComtrerazAún no hay calificaciones
- PROPIEDADES ELÉCTRICAS DEL SISTEMA NERVIOSO - Notas de ClaseDocumento10 páginasPROPIEDADES ELÉCTRICAS DEL SISTEMA NERVIOSO - Notas de ClasewpmariaantoniaAún no hay calificaciones
- Comunicación NerviosaDocumento3 páginasComunicación NerviosaG Priscila P. IpertyAún no hay calificaciones
- Bases de ElectrofisiologíaDocumento12 páginasBases de ElectrofisiologíaDAYELY NICOLL DELGADO ROJASAún no hay calificaciones
- Tema 10 SINAPSIS Imp PDFDocumento27 páginasTema 10 SINAPSIS Imp PDFMar DíazAún no hay calificaciones
- Informe Fisiologia - Power LabDocumento11 páginasInforme Fisiologia - Power LabFelipe SanchezAún no hay calificaciones
- Semiologia Ocular 2006Documento102 páginasSemiologia Ocular 2006Ronald Torres ChávezAún no hay calificaciones
- FisiologíaDocumento24 páginasFisiologíajoaquinAún no hay calificaciones
- Condicionamiento Operante y ClasicoDocumento33 páginasCondicionamiento Operante y ClasicoJulioAdalbertoAún no hay calificaciones
- Clase 2 Sinapsis y Contracción Musculae 2023Documento26 páginasClase 2 Sinapsis y Contracción Musculae 2023km100% (1)
- Capitulo Señalizacion Celular-ReceptoresDocumento60 páginasCapitulo Señalizacion Celular-ReceptoresEdwar Leyva Baltazar0% (1)
- Potenciales de Accion Motor Y Sensorial: Manifestaciones Neurológicas de La Lesión Nerviosa Y Su CorrelaciónDocumento61 páginasPotenciales de Accion Motor Y Sensorial: Manifestaciones Neurológicas de La Lesión Nerviosa Y Su Correlaciónjonesc9696Aún no hay calificaciones
- Modulo Profesonal de Soporte TecnicoDocumento54 páginasModulo Profesonal de Soporte TecnicoLuciana VicenteAún no hay calificaciones
- Cuestionario 5 Luis Cabrera Vertiz 03 SetiembreDocumento6 páginasCuestionario 5 Luis Cabrera Vertiz 03 SetiembreRogerHenryAguirreTrujilloAún no hay calificaciones
- Ello, Yo y SuperyóDocumento6 páginasEllo, Yo y SuperyóJohnAún no hay calificaciones
- Resumen Neurobiología FINALDocumento98 páginasResumen Neurobiología FINALCynthia ScottiAún no hay calificaciones
- Electro Encefalograma EspontaneoDocumento4 páginasElectro Encefalograma EspontaneoMaFeerly ZuñiGaAún no hay calificaciones
- 2.1 Tema-EncefalogramaDocumento43 páginas2.1 Tema-Encefalogramacarlota100% (1)
- Estudio Biofisico de Los Canales IonicosDocumento8 páginasEstudio Biofisico de Los Canales IonicospedroAún no hay calificaciones
- 1er PARCIAL FISIOLOGIA NEURO PDFDocumento37 páginas1er PARCIAL FISIOLOGIA NEURO PDFJsgm24Aún no hay calificaciones
- Actualizar BiosDocumento10 páginasActualizar BiosGlory VIlcaAún no hay calificaciones
- Ablación ExperimentalDocumento3 páginasAblación ExperimentalMary VgAún no hay calificaciones
- Sistema Somatosensorial 6Documento29 páginasSistema Somatosensorial 6Rafael DazaAún no hay calificaciones
- Técnica Patch ClampDocumento3 páginasTécnica Patch ClampAlejandroHidalgoNavaAún no hay calificaciones
- Que Son Las Neuroglias y Su FuncionDocumento5 páginasQue Son Las Neuroglias y Su Funcionnelys lopezAún no hay calificaciones
- Practica 1 Antenas y PropagacionDocumento17 páginasPractica 1 Antenas y PropagacionGeenaTatianaMalaverPrietoAún no hay calificaciones
- Fibra Optica SubmarinaDocumento2 páginasFibra Optica SubmarinaFrank Pacherre MoralesAún no hay calificaciones
- Neurofisiologia I y IIDocumento157 páginasNeurofisiologia I y IIUSMP FN ARCHIVOSAún no hay calificaciones
- Guía N6 Neurona Estru y FunciónDocumento15 páginasGuía N6 Neurona Estru y FunciónNico Ignacio VeraAún no hay calificaciones
- Potenciales de Membrana y Potenciales de AcciónDocumento31 páginasPotenciales de Membrana y Potenciales de AcciónAnto FloresAún no hay calificaciones
- Resúmen Neurofisiología (1er Parcial)Documento20 páginasResúmen Neurofisiología (1er Parcial)Mili NunesAún no hay calificaciones
- Capitulo 55Documento5 páginasCapitulo 55leslie laraAún no hay calificaciones
- HormonasDocumento15 páginasHormonasJhon Kevin Bonilla LeónAún no hay calificaciones
- Apoptosis Celular PDFDocumento4 páginasApoptosis Celular PDFJhonny Rolando ConchaAún no hay calificaciones
- Estructura y Funcion Del Sistema NerviosoDocumento23 páginasEstructura y Funcion Del Sistema NerviosoXiomy RmAún no hay calificaciones
- Función Nerviosa de La RetinaDocumento5 páginasFunción Nerviosa de La RetinaDAICY EDITH LEDEZMA PEREZAún no hay calificaciones
- Resumen. ExcitabilidadDocumento39 páginasResumen. ExcitabilidadMiguelangel VeraAún no hay calificaciones
- NeuronasDocumento10 páginasNeuronasGadeli Ventura MontoyaAún no hay calificaciones
- Tema 11. A. Técnicas de NeuroimagenDocumento46 páginasTema 11. A. Técnicas de NeuroimagenRCRAún no hay calificaciones
- El Potencial de Acción o Potencial de MembranaDocumento6 páginasEl Potencial de Acción o Potencial de MembranaJENNIFER DANIELA MARTINEZ LOPEZAún no hay calificaciones
- Neurona, Potencial de Accion y ReposoDocumento13 páginasNeurona, Potencial de Accion y ReposoluyhlodelaAún no hay calificaciones
- Edelman Tononi El Universo de La Conciencia Cap 5-6-13Documento33 páginasEdelman Tononi El Universo de La Conciencia Cap 5-6-13Carlos Roa100% (1)
- 3 Vias Espinales - m11wDocumento12 páginas3 Vias Espinales - m11wLeticia Verdugo Hernandez She HerAún no hay calificaciones
- Sistema de EndomembranasDocumento41 páginasSistema de EndomembranasTomas DelgadoAún no hay calificaciones
- Silabo Salud Mental ESPOCHDocumento32 páginasSilabo Salud Mental ESPOCHKarelys AndradeAún no hay calificaciones
- Neurociencias Clase I Neuronas Circuitos Neuronales y ConductaDocumento19 páginasNeurociencias Clase I Neuronas Circuitos Neuronales y ConductaMacarena CepedaAún no hay calificaciones
- Mini Ebook Neuroanatomia 83269 Downloable 1270085Documento143 páginasMini Ebook Neuroanatomia 83269 Downloable 1270085Dennís LlacctarimayAún no hay calificaciones
- Unidad II (1b) Regla Hebb - NntoolDocumento25 páginasUnidad II (1b) Regla Hebb - NntoolTomás Andrés Madrid RosalesAún no hay calificaciones
- Presentacion IoT PDFDocumento35 páginasPresentacion IoT PDFkeneth portillaAún no hay calificaciones
- Tarea 1 - Instrumentacion MedicaDocumento11 páginasTarea 1 - Instrumentacion MedicaJulian Andres Hernandez CastroAún no hay calificaciones
- BiopotencialesDocumento241 páginasBiopotencialesHéctor Alan AguilarAún no hay calificaciones
- BiopotencialesDocumento29 páginasBiopotencialesYakeline VelasquezAún no hay calificaciones
- CARTILLA UNAL Bogota BiologiaDocumento19 páginasCARTILLA UNAL Bogota BiologiaJuan DavidAún no hay calificaciones
- El Origen Del Universo Según Los EgipciosDocumento2 páginasEl Origen Del Universo Según Los EgipciosLourdes BadilloAún no hay calificaciones
- MICORRIZASDocumento8 páginasMICORRIZASjosephAún no hay calificaciones
- Tercer Parcial de BioquimicaDocumento20 páginasTercer Parcial de BioquimicaJOSE MANUEL MORILLO100% (1)
- Trabajo 1 - Ing. de Recursos Hidraulicos PDFDocumento119 páginasTrabajo 1 - Ing. de Recursos Hidraulicos PDFMarco DanielAún no hay calificaciones
- Horario Joaquin 2024Documento2 páginasHorario Joaquin 2024hamburgesa GAMERAún no hay calificaciones
- 20 Preguntas de Músculo EstructuraDocumento2 páginas20 Preguntas de Músculo EstructuraNéstor Raúl Henao SierraAún no hay calificaciones
- Indice Libro IberáDocumento15 páginasIndice Libro IberáVeronicaAún no hay calificaciones
- Insecticida NaturalDocumento6 páginasInsecticida NaturalCami TravieAún no hay calificaciones
- Cambiop de Giro de Spool Tipo Yee y Tee MetsoDocumento27 páginasCambiop de Giro de Spool Tipo Yee y Tee MetsoJavier Serpa Quispe100% (1)
- 8 Peores Sismos en Peru - Sismos 1Documento31 páginas8 Peores Sismos en Peru - Sismos 1Jorge Luis Enrique SalazarAún no hay calificaciones
- Comparación de Métodos de Recolección de Gases (Por Pérdida de Masa y Desplazamiento de Agua)Documento7 páginasComparación de Métodos de Recolección de Gases (Por Pérdida de Masa y Desplazamiento de Agua)erbrow2075% (4)
- VENTANA A MI COMUNIDAD EL PUEBL - Luz Maria Chapela PDFDocumento56 páginasVENTANA A MI COMUNIDAD EL PUEBL - Luz Maria Chapela PDFRigoberto Vicente CastroAún no hay calificaciones
- RV - Textos - Iv (Teoría - Práctica)Documento9 páginasRV - Textos - Iv (Teoría - Práctica)luisangeluni100% (1)
- Cancionero ReggaeDocumento25 páginasCancionero ReggaeVitoko Espinoza VivarAún no hay calificaciones
- Secciones Cónicas (Elipse)Documento24 páginasSecciones Cónicas (Elipse)Luis CarlosAún no hay calificaciones
- G2 Genetic AlgorithmsDocumento58 páginasG2 Genetic AlgorithmsNoe Cuellar MartinezAún no hay calificaciones
- Caja de Herramientas - Dinamicas FormativasDocumento307 páginasCaja de Herramientas - Dinamicas FormativasYULI JEJENAún no hay calificaciones
- Los Origenes de Los PecesDocumento20 páginasLos Origenes de Los PecesBetty D'EmidioAún no hay calificaciones
- Revista DeverasDocumento2 páginasRevista DeverasJorgeBecerraAún no hay calificaciones
- Ficha Genero Del Sustantivo para Tercero de PrimariaDocumento3 páginasFicha Genero Del Sustantivo para Tercero de PrimariaLidya Cardenas67% (6)
- EVALUACIÓNDocumento2 páginasEVALUACIÓNNoes Pi100% (1)
- Pasteur y KochDocumento2 páginasPasteur y KochJavier MartínezAún no hay calificaciones
- Anteproyecto de ResidenciasDocumento11 páginasAnteproyecto de ResidenciasSandra Luz Cruz HernandezAún no hay calificaciones
- Análisis de Aptitud para La Expansión UrbanaDocumento1 páginaAnálisis de Aptitud para La Expansión UrbanaKate RojasAún no hay calificaciones
- Cartilla PPPDocumento36 páginasCartilla PPPAlberto Lozada SilvaAún no hay calificaciones
- Metodologia de La Investigacion II. Tarea 2Documento8 páginasMetodologia de La Investigacion II. Tarea 2JeuryAún no hay calificaciones