También podría gustarte
- Cuestionario Urinario PDFDocumento6 páginasCuestionario Urinario PDFDANIEL EDUARDO VALDEZ ORTEGAAún no hay calificaciones
- Características Del Reino MetazoaDocumento9 páginasCaracterísticas Del Reino MetazoaGerardo SandovalAún no hay calificaciones
- Informe 3Documento13 páginasInforme 3Jose Luis Condori PintoAún no hay calificaciones
- Reporte de Práctica de Artrópodos No InsectosDocumento11 páginasReporte de Práctica de Artrópodos No InsectosPerla Quiñonez QuevedoAún no hay calificaciones
- Mazariegos Emilio - Salmos de Un Corazon JovenDocumento115 páginasMazariegos Emilio - Salmos de Un Corazon JovenGloria Elena Perez Morales86% (14)
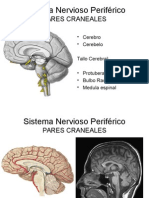
- Pares Craneales - Es Slideshare Net 34Documento34 páginasPares Craneales - Es Slideshare Net 34Felipe Riestra0% (1)
- Tarea de Craneo Grupo 2Documento26 páginasTarea de Craneo Grupo 2Sugeily CastilloAún no hay calificaciones
- Bioqumica Ciclo de KrebsDocumento9 páginasBioqumica Ciclo de KrebsTatiana de los Ángeles López PérezAún no hay calificaciones
- CuestionarioDocumento3 páginasCuestionarioLuis CossaAún no hay calificaciones
- Intoxicación Por Sulfato de CobreDocumento23 páginasIntoxicación Por Sulfato de CobreJesus Lopez Hernandez50% (2)
- Saladin Anatomia 6a Diapositivas c24 HIDROELECTROLITICODocumento14 páginasSaladin Anatomia 6a Diapositivas c24 HIDROELECTROLITICOFranklin garryAún no hay calificaciones
- Proyecto de Morfoanatomia Vegetal (La Hoja)Documento10 páginasProyecto de Morfoanatomia Vegetal (La Hoja)Jhonatan Lara LópezAún no hay calificaciones
- Ejercicio 1 EnzimologiaDocumento15 páginasEjercicio 1 EnzimologiaAndrea Chavarro100% (1)
- Propiedades Del LicopenoDocumento2 páginasPropiedades Del LicopenoKevin Steven Asto RomeroAún no hay calificaciones
- Reseña Diez TesisDocumento5 páginasReseña Diez TesisHernán CanoAún no hay calificaciones
- 3 CelulaDocumento39 páginas3 CelulaAriadna Wolf100% (1)
- MetabolismoDocumento4 páginasMetabolismoArévaloAún no hay calificaciones
- Kawauchi2014 TraducidoDocumento26 páginasKawauchi2014 TraducidoSofia CantúAún no hay calificaciones
- Reptiles PDFDocumento35 páginasReptiles PDFalegarma19Aún no hay calificaciones
- FitohemaglutininaDocumento9 páginasFitohemaglutininaChristina ZtaAún no hay calificaciones
- Aminoácidos No Proteicos en La Defensa de Plantas Contra Herbívoros de InsectosDocumento10 páginasAminoácidos No Proteicos en La Defensa de Plantas Contra Herbívoros de InsectosLizeth León CarreñoAún no hay calificaciones
- ProteinasDocumento6 páginasProteinasNataliaAún no hay calificaciones
- Importancia de Las ProteinasDocumento3 páginasImportancia de Las ProteinasNicoleAún no hay calificaciones
- Los Organismos y Su AmbienteDocumento3 páginasLos Organismos y Su AmbienteMhi-mi Thi-fany Q CAún no hay calificaciones
- Teoría Del Mundo de HierroDocumento2 páginasTeoría Del Mundo de HierroJeeniffer Cookiee100% (2)
- Vitamina B2Documento3 páginasVitamina B2Josselyn RodrguezAún no hay calificaciones
- Rally PDFDocumento10 páginasRally PDFDany CamposAún no hay calificaciones
- Clasificación BacterianaDocumento12 páginasClasificación BacterianaElsi Mariann Gutierrez GarciaAún no hay calificaciones
- Linus PaulingDocumento18 páginasLinus Paulingjovmac2uAún no hay calificaciones
- REINOS DE LA NATURALEZA Cesar RodriguezDocumento3 páginasREINOS DE LA NATURALEZA Cesar Rodriguezwalter martinezAún no hay calificaciones
- Guía de Biología 2º Medio Hormonas PrincipalesDocumento9 páginasGuía de Biología 2º Medio Hormonas Principalescienciasaltazor100% (3)
- 2 IntroduccionDocumento36 páginas2 IntroduccionNorma VillalvaAún no hay calificaciones
- Obtencion de AmilasaDocumento16 páginasObtencion de Amilasadalfredo018Aún no hay calificaciones
- Lectura 1 BQ PDFDocumento4 páginasLectura 1 BQ PDFAlexiss Chavez-De la MoraAún no hay calificaciones
- BiologiaDocumento15 páginasBiologiaMichael Gutiérrez BenitezAún no hay calificaciones
- Síntesis de NucleótidosDocumento3 páginasSíntesis de NucleótidosAnonymous guhIUozAún no hay calificaciones
- Guiaexcrecion 2011Documento7 páginasGuiaexcrecion 2011glagarbe271100% (1)
- Evolucion de Los ParasitosDocumento1 páginaEvolucion de Los ParasitosFidel CortesAún no hay calificaciones
- OsmoralidadDocumento7 páginasOsmoralidadMARIA XIMENA ZARATE JEREZAún no hay calificaciones
- ATEROGENESISDocumento14 páginasATEROGENESISLiz Matamoros MezaAún no hay calificaciones
- LIDERAZGO (Biologí A y Sociedad) : Área 9. Ciencias Sociales y Económico AdministrativasDocumento213 páginasLIDERAZGO (Biologí A y Sociedad) : Área 9. Ciencias Sociales y Económico AdministrativasK A R L A M U R I L L OAún no hay calificaciones
- Exam Resuelto de Biolo 1xpuntoDocumento4 páginasExam Resuelto de Biolo 1xpuntoWilly BaldelomarAún no hay calificaciones
- Manual de Prácticas de FisiologíaDocumento147 páginasManual de Prácticas de FisiologíaJessy JessyAún no hay calificaciones
- Actividad EnzimaticaDocumento7 páginasActividad EnzimaticaKristel Alexandra Lopez RamirezAún no hay calificaciones
- Guia PHDocumento11 páginasGuia PHLiliana CupitraAún no hay calificaciones
- Zoologia GD 1bt 2022 ADocumento126 páginasZoologia GD 1bt 2022 APepe GaonAún no hay calificaciones
- SpirulinaDocumento8 páginasSpirulinaDiana MaciasAún no hay calificaciones
- Arqueas-Y-Bacterias Trabajo MonograficoDocumento26 páginasArqueas-Y-Bacterias Trabajo MonograficoAntonio Ulloa TorresAún no hay calificaciones
- Gasparin - Líquidos BiológicosDocumento5 páginasGasparin - Líquidos BiológicosJOSELYN MISHELL ARAUZ SIERRAAún no hay calificaciones
- Phylum HemicordadosDocumento15 páginasPhylum Hemicordadosyanina sosa100% (1)
- Panthera Onca PDFDocumento51 páginasPanthera Onca PDFalejandro pazAún no hay calificaciones
- GeranioDocumento3 páginasGeranioAlondra NavaAún no hay calificaciones
- Que Es La MicrodosisDocumento3 páginasQue Es La MicrodosisSantos CotoAún no hay calificaciones
- Exposicion MesozoicoDocumento2 páginasExposicion MesozoicoExdiliany RevetteAún no hay calificaciones
- Acumulaciones IntracelularesDocumento3 páginasAcumulaciones IntracelularesYeliLagunaAún no hay calificaciones
- Taller 4 EnzimasDocumento2 páginasTaller 4 EnzimasJegase Kibo33% (3)
- Biocelular (CAP.6) FOTOSÍNTESIS, LUZ Y VIDA-PAG-145-164Documento20 páginasBiocelular (CAP.6) FOTOSÍNTESIS, LUZ Y VIDA-PAG-145-164Mishell Daffne Mendoza FloresAún no hay calificaciones
- Funciones Membrana PlasmaticaDocumento9 páginasFunciones Membrana PlasmaticaJossué LataAún no hay calificaciones
- Tp1-Quimicabiologica2-Sinthia Lucrecia AguireDocumento5 páginasTp1-Quimicabiologica2-Sinthia Lucrecia AguireLucrecia AguirreAún no hay calificaciones
- Transporte A Traves Demembrana Celular 5Documento22 páginasTransporte A Traves Demembrana Celular 5Jhon Alexis M ArgoteAún no hay calificaciones
- Mhs Tramo Dos AlafDocumento59 páginasMhs Tramo Dos AlafZiaraAún no hay calificaciones
- Conductos Y Bombas de Membrana: Escuela Profesional de Biologia-Bioquimica IiDocumento63 páginasConductos Y Bombas de Membrana: Escuela Profesional de Biologia-Bioquimica IiClinton LunaAún no hay calificaciones
- Informe de Trabajos de Mantenimiento Julio 2020Documento11 páginasInforme de Trabajos de Mantenimiento Julio 2020Börys PalmaAún no hay calificaciones
- Pc1 Caf3 2021 OficialDocumento45 páginasPc1 Caf3 2021 OficialRichar Mario Delgado ChiloAún no hay calificaciones
- Guía 2 Clasificación Materia Átomos MolDocumento2 páginasGuía 2 Clasificación Materia Átomos MolJosé ArévaloAún no hay calificaciones
- Alcoholismo en El Mundo - AnatomiaDocumento6 páginasAlcoholismo en El Mundo - AnatomiafreddyelmenAún no hay calificaciones
- Discusio 2 AnaliticaDocumento3 páginasDiscusio 2 AnaliticaLuis SigüenzaAún no hay calificaciones
- Ensayo La CrisisDocumento5 páginasEnsayo La CrisissandraAún no hay calificaciones
- Pts Camion RRDDocumento7 páginasPts Camion RRDPABLO ANDRES PEREIRA SILVAAún no hay calificaciones
- Metabolismo CardiacoDocumento14 páginasMetabolismo CardiacoAndrés León AristizábalAún no hay calificaciones
- Diagnostico de Salud ComunitarioDocumento27 páginasDiagnostico de Salud ComunitarioGiio NoocheebueenaaAún no hay calificaciones
- Proyecto FinalDocumento21 páginasProyecto FinalJOHANA JANETH JARABA DEDEAún no hay calificaciones
- 16 Partes Que Conforman La Célula VegetalDocumento3 páginas16 Partes Que Conforman La Célula VegetalCarlos JuárezAún no hay calificaciones
- Experimentos Caseros para NiNos IIDescubriendo El AguaDocumento59 páginasExperimentos Caseros para NiNos IIDescubriendo El AguaMónica CairoliAún no hay calificaciones
- Caracteristicas Climaticas de Las Regiones VenezolanasDocumento13 páginasCaracteristicas Climaticas de Las Regiones VenezolanasGisela Lopez50% (4)
- Info TUTORIADocumento2 páginasInfo TUTORIASegundo MarinAún no hay calificaciones
- Invitación AFA COLOMBIADocumento3 páginasInvitación AFA COLOMBIACARLOS ALBERTO DIAZ QUIROZAún no hay calificaciones
- Práctica de MezclasDocumento4 páginasPráctica de MezclasJuan Pablo Rodriguez ZamoraAún no hay calificaciones
- Trans Mision SexualDocumento3 páginasTrans Mision SexualfrancesubvAún no hay calificaciones
- Grupo Iii Abc 5° 2017Documento4 páginasGrupo Iii Abc 5° 2017AntonioIslaTapiaAún no hay calificaciones
- Taller Postrados Cesfam Sta AmaliaDocumento99 páginasTaller Postrados Cesfam Sta AmaliaPatricio Pino Mora0% (1)
- MR Folleto Caudalimetro Electromagnetico de Inserción MR-INa Y MR-INbDocumento3 páginasMR Folleto Caudalimetro Electromagnetico de Inserción MR-INa Y MR-INbVladimir Laura DelgadoAún no hay calificaciones
- P1-Material de Uso Comun en El Laboratorio, Segurida e HigieneDocumento9 páginasP1-Material de Uso Comun en El Laboratorio, Segurida e HigienePancracia GarciaAún no hay calificaciones
- Basculante 1Documento109 páginasBasculante 1Angel Andrade GarciaAún no hay calificaciones
- Cromatografia de Capa DelgadaDocumento22 páginasCromatografia de Capa DelgadaJeremy Nicolás León CamposAún no hay calificaciones
- A Review of Selected Dental Literature On Conteemporary Provisional - En.esDocumento24 páginasA Review of Selected Dental Literature On Conteemporary Provisional - En.esMartha Gabriela SanchezAún no hay calificaciones
- Manual PSB Limpieza y Desinfección FinalDocumento17 páginasManual PSB Limpieza y Desinfección FinalIZKETA "Segovia" INSTITUCION ETNOEDUCATIVA U'WAAún no hay calificaciones
- Guía Dia Del Idioma 2023Documento4 páginasGuía Dia Del Idioma 2023Sonia Lucero García BlancoAún no hay calificaciones
- Sindrome de BehrDocumento3 páginasSindrome de Behrapi-3697492Aún no hay calificaciones
- IV FIN 110 TE Chura Anticona 2019Documento101 páginasIV FIN 110 TE Chura Anticona 2019Jean Claudio LazzarteAún no hay calificaciones