100% encontró este documento útil (3 votos)
582 vistas96 páginasBioelectricidad y Transporte Iónico Celular
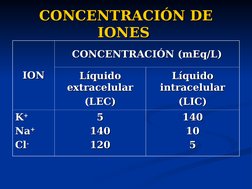
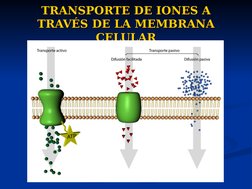
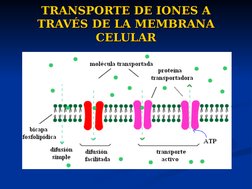
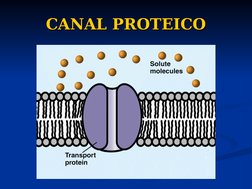
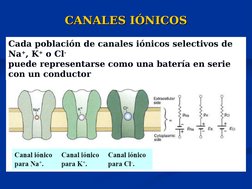
El documento describe conceptos básicos de bioelectromagnetismo, incluyendo la conducción de corrientes iónicas en organismos vivos, la polarización de la membrana celular, el transporte de iones a través de la membrana mediante difusión pasiva, difusión facilitada y transporte activo, y el potencial de reposo de las células. También explica conceptos clave como el potencial de acción, las ecuaciones de Nernst y Goldman, y los elementos eléctricos equivalentes de la célula nerviosa.
Cargado por
Fátima Arias CabrejosDerechos de autor
© © All Rights Reserved
Nos tomamos en serio los derechos de los contenidos. Si sospechas que se trata de tu contenido, reclámalo aquí.
Formatos disponibles
Descarga como PPT, PDF, TXT o lee en línea desde Scribd
100% encontró este documento útil (3 votos)
582 vistas96 páginasBioelectricidad y Transporte Iónico Celular
El documento describe conceptos básicos de bioelectromagnetismo, incluyendo la conducción de corrientes iónicas en organismos vivos, la polarización de la membrana celular, el transporte de iones a través de la membrana mediante difusión pasiva, difusión facilitada y transporte activo, y el potencial de reposo de las células. También explica conceptos clave como el potencial de acción, las ecuaciones de Nernst y Goldman, y los elementos eléctricos equivalentes de la célula nerviosa.
Cargado por
Fátima Arias CabrejosDerechos de autor
© © All Rights Reserved
Nos tomamos en serio los derechos de los contenidos. Si sospechas que se trata de tu contenido, reclámalo aquí.
Formatos disponibles
Descarga como PPT, PDF, TXT o lee en línea desde Scribd