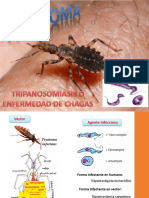
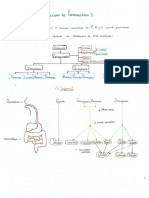
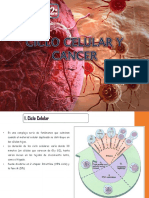
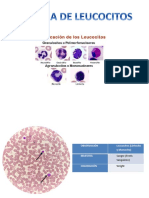
También podría gustarte
- Caso Clinico Motor Semana VDocumento2 páginasCaso Clinico Motor Semana VJr EscobarAún no hay calificaciones
- ANTIMICROBIANOSDocumento4 páginasANTIMICROBIANOSJr EscobarAún no hay calificaciones
- Expo SaludDocumento12 páginasExpo SaludJr EscobarAún no hay calificaciones
- 20190913180926Documento1 página20190913180926Jr EscobarAún no hay calificaciones
- Genetica de Enfermedad HDocumento22 páginasGenetica de Enfermedad HJr EscobarAún no hay calificaciones
- Tejido Conectivo Histo 4Documento102 páginasTejido Conectivo Histo 4Jr EscobarAún no hay calificaciones
- Inf GenéticaDocumento5 páginasInf GenéticaJr EscobarAún no hay calificaciones
- InformeDocumento1 páginaInformeJr EscobarAún no hay calificaciones
- Resumen de Articulo FisioDocumento2 páginasResumen de Articulo FisioJr EscobarAún no hay calificaciones
- Genetica SILABODocumento9 páginasGenetica SILABOAndreaAún no hay calificaciones
- eJ.J.A: G (A, R - B. "Documento3 páginaseJ.J.A: G (A, R - B. "Jr EscobarAún no hay calificaciones
- SeminarioDocumento57 páginasSeminarioJr EscobarAún no hay calificaciones
- HistoDocumento5 páginasHistoJr EscobarAún no hay calificaciones
- Seminario GehiemDocumento18 páginasSeminario GehiemJr EscobarAún no hay calificaciones
- Membrana PlasmáticaDocumento20 páginasMembrana PlasmáticaJr EscobarAún no hay calificaciones
- Mastocitos GenéticaDocumento3 páginasMastocitos GenéticaJr EscobarAún no hay calificaciones
- Practica 2-Lipidos y ProteínasDocumento19 páginasPractica 2-Lipidos y ProteínasJr EscobarAún no hay calificaciones
- DIA 7 y 8 EmbrioDocumento2 páginasDIA 7 y 8 EmbrioJr EscobarAún no hay calificaciones
- PleiotropíaDocumento1 páginaPleiotropíaJr EscobarAún no hay calificaciones
- Genética e Histoembriología: Observación Muestra Coloración AumentoDocumento1 páginaGenética e Histoembriología: Observación Muestra Coloración AumentoJr EscobarAún no hay calificaciones
- Seminario IIDocumento67 páginasSeminario IIJr EscobarAún no hay calificaciones
- Seminario IIDocumento67 páginasSeminario IIJr EscobarAún no hay calificaciones
- Monografia BiofisicaDocumento20 páginasMonografia BiofisicaJr EscobarAún no hay calificaciones
- InformaciónDocumento6 páginasInformaciónJr EscobarAún no hay calificaciones
- Ex PosicionDocumento45 páginasEx PosicionJr EscobarAún no hay calificaciones
- Requisitos Creditos PYMEDocumento1 páginaRequisitos Creditos PYMEJr EscobarAún no hay calificaciones
- EXPOSICIONDocumento9 páginasEXPOSICIONJr EscobarAún no hay calificaciones
- CpanelDocumento30 páginasCpanelJr EscobarAún no hay calificaciones
- Transporte A Través de La Membrana PlasmáticaDocumento17 páginasTransporte A Través de La Membrana PlasmáticaCarola DJAún no hay calificaciones
- Neuro Transm I SoresDocumento38 páginasNeuro Transm I SoresDANIEL STANLEY VENTURA ROQUEAún no hay calificaciones
- Articulo 1 ESDocumento21 páginasArticulo 1 ESAndrésAún no hay calificaciones
- Tema 8. B. Audesirk 2013-Nerv 734-743Documento11 páginasTema 8. B. Audesirk 2013-Nerv 734-743Marco Joel Pinto MaidanaAún no hay calificaciones
- 15 - Sistema de Conduccion, Vectores y Ondas 2022Documento64 páginas15 - Sistema de Conduccion, Vectores y Ondas 2022CarlosAún no hay calificaciones
- Farmacocinetica y FarmacodinamiaDocumento8 páginasFarmacocinetica y FarmacodinamiaGabriela CárdenasAún no hay calificaciones
- Tolerancia de Los Cultivos Al Estrés SalinoDocumento22 páginasTolerancia de Los Cultivos Al Estrés SalinoAbi CárdenasAún no hay calificaciones
- Farmacologia Clase 3 FinDocumento31 páginasFarmacologia Clase 3 FinCatalina MarinAún no hay calificaciones
- Impulso Nervioso, Sinapsis y NeurotransmisoresDocumento17 páginasImpulso Nervioso, Sinapsis y NeurotransmisoresDagoberto Andres Fuentes Solis100% (2)
- Clase 2-Fisiologia NeuronalDocumento11 páginasClase 2-Fisiologia NeuronalkasikeAún no hay calificaciones
- Resumen de Biología CelularDocumento67 páginasResumen de Biología CelularSofia DuarteAún no hay calificaciones
- Farmacos de AnestesiaDocumento57 páginasFarmacos de AnestesiaPaula Zambrano100% (1)
- Compendio FisioDocumento278 páginasCompendio FisiobjotaleAún no hay calificaciones
- Práctica 3 "Simulación Por Computadora Del Potencial de Acción"Documento24 páginasPráctica 3 "Simulación Por Computadora Del Potencial de Acción"Diego Vel100% (1)
- Fisiología Del Sistema NerviosoDocumento78 páginasFisiología Del Sistema NerviosoEmiliano A. DorelleAún no hay calificaciones
- ElectrocardiografiaDocumento33 páginasElectrocardiografiaMonica Garcia AiquipaAún no hay calificaciones
- Receptores FarmacologicosDocumento76 páginasReceptores FarmacologicosPedro José Ortíz Ortega100% (2)
- Potencial de Acción CardiacoDocumento13 páginasPotencial de Acción CardiacoMenelo Soliman RodriguezAún no hay calificaciones
- Resumen 2do CuatrimestreDocumento132 páginasResumen 2do CuatrimestrevaleAún no hay calificaciones
- Copia de Copia de Osuna Cap 1Documento39 páginasCopia de Copia de Osuna Cap 1Karen ArciniegasAún no hay calificaciones
- Compendio de Anestesiolog°a 2014 PDFDocumento162 páginasCompendio de Anestesiolog°a 2014 PDFJenaro AlvarezAún no hay calificaciones
- Cap 7 Fármacos Activadores de Colinorreceptores e Inhibidores de ColinesterasaDocumento10 páginasCap 7 Fármacos Activadores de Colinorreceptores e Inhibidores de Colinesterasarodrigo4590100% (1)
- Libro Resumen: Por Estudiantes Medicina de La Universidad Peruana Cayetano HerediaDocumento69 páginasLibro Resumen: Por Estudiantes Medicina de La Universidad Peruana Cayetano HerediaAlejandra Llerena100% (1)
- Practica 3 FisiologiaDocumento14 páginasPractica 3 FisiologiaLuisa RVAún no hay calificaciones
- Guía de Práctica de Laboratorio de 11111111111111111Documento19 páginasGuía de Práctica de Laboratorio de 11111111111111111JESSELY AMPARO FLORES ALVAREZAún no hay calificaciones
- Clase 2 - Principios Generales de La Acción de Los FármacosDocumento59 páginasClase 2 - Principios Generales de La Acción de Los FármacosJose AcostaAún no hay calificaciones
- Neuroquimica y NeurotransmisoresDocumento62 páginasNeuroquimica y NeurotransmisoresdanAún no hay calificaciones
- Señalizacion CelularDocumento60 páginasSeñalizacion CelularcarlosAún no hay calificaciones
- Psicofarmacos Primer Parcial Semanas 1 A 6Documento251 páginasPsicofarmacos Primer Parcial Semanas 1 A 6Facundo LopezAún no hay calificaciones
- 1 Termodinámica Potencial Químico y ElectroquímicoDocumento7 páginas1 Termodinámica Potencial Químico y ElectroquímicoDennise San MartinAún no hay calificaciones