También podría gustarte
- Clasificacion Cernea y OtrosDocumento8 páginasClasificacion Cernea y OtrosKevin Lois Cesías MeléndezAún no hay calificaciones
- Resultados-MARIA ANGELA PANCA DIAZDocumento1 páginaResultados-MARIA ANGELA PANCA DIAZKevin Lois Cesías MeléndezAún no hay calificaciones
- Colgajos Cutáneos en La Reconstrucción de La Región Supra y ParaciliarDocumento10 páginasColgajos Cutáneos en La Reconstrucción de La Región Supra y ParaciliarKevin Lois Cesías MeléndezAún no hay calificaciones
- Infecciones Orofaciales Cabeza Cuello (Hupp y Ferneini)Documento495 páginasInfecciones Orofaciales Cabeza Cuello (Hupp y Ferneini)Kevin Lois Cesías Meléndez100% (1)
- Acto MedicoDocumento30 páginasActo MedicoKevin Lois Cesías MeléndezAún no hay calificaciones
- NT 0021 Documento Oficial CategorizacionDocumento91 páginasNT 0021 Documento Oficial CategorizacionCarl0zzAún no hay calificaciones
- OligoelementosDocumento9 páginasOligoelementosilamacamachojesusAún no hay calificaciones
- Unidad 3 y 4 Plan de Vida y Carrera Profesional.Documento5 páginasUnidad 3 y 4 Plan de Vida y Carrera Profesional.Leez Ontiveros DiazAún no hay calificaciones
- 1T-Introduccion A ReproDocumento19 páginas1T-Introduccion A ReproJose Emma Tamo CasaAún no hay calificaciones
- Prueba Coeficiente 2 de Ciencias Naturales FinalDocumento2 páginasPrueba Coeficiente 2 de Ciencias Naturales FinalDavid Zamora JeldesAún no hay calificaciones
- 3-Tema Desarrollo de Aptitud, Conocimiento y CapacidadDocumento22 páginas3-Tema Desarrollo de Aptitud, Conocimiento y CapacidadGuillermo Enrique Rivas OteroAún no hay calificaciones
- Biometria Fetal2Documento120 páginasBiometria Fetal2Luis Medina CoriaAún no hay calificaciones
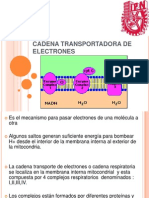
- Cadena Transport Ad or A de ElectronesDocumento17 páginasCadena Transport Ad or A de Electronesjimmy_caolAún no hay calificaciones
- La Vida de Los PecesDocumento5 páginasLa Vida de Los PecesPaula TedeschiAún no hay calificaciones
- Periodoncia I 2 OrishinalDocumento35 páginasPeriodoncia I 2 OrishinalNicolasAún no hay calificaciones
- EritrocitoDocumento6 páginasEritrocitoDayana AguilarAún no hay calificaciones
- XS 1000i EspañolDocumento8 páginasXS 1000i EspañolJimmy Rodríguez JuárezAún no hay calificaciones
- Laboratorio Diseccion de CorazonDocumento3 páginasLaboratorio Diseccion de CorazonDiego Alejandro AlvaranAún no hay calificaciones
- DentalDocumento6 páginasDentalAngel Gonzalez ArmendarizAún no hay calificaciones
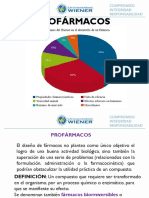
- FarmacoquimicaDocumento28 páginasFarmacoquimicaOscar Paredes0% (1)
- 14 Caso Clinico FemeninoDocumento2 páginas14 Caso Clinico FemeninoEstefany PérezAún no hay calificaciones
- Inmunocromatografia de VIHDocumento8 páginasInmunocromatografia de VIHEdgarAún no hay calificaciones
- CARDIOPATIADocumento25 páginasCARDIOPATIADanielaAún no hay calificaciones
- Rubrica para Evaluar Portafolio de Evidencias 2016 n300Documento6 páginasRubrica para Evaluar Portafolio de Evidencias 2016 n300ALEJANDRO OSTOIC ROZZIAún no hay calificaciones
- PDF 3Documento3 páginasPDF 3karen DextreAún no hay calificaciones
- Arritmias LetalesDocumento9 páginasArritmias LetalesArteAún no hay calificaciones
- Andreas Ludwig Kalcker - ParásitosDocumento13 páginasAndreas Ludwig Kalcker - ParásitosAnonymous MXBabgkV100% (1)
- Gluconeogenesis PPT YesiDocumento20 páginasGluconeogenesis PPT YesiCesar Sarmiento PerezAún no hay calificaciones
- Biologia 5Documento12 páginasBiologia 5Alejandra Martin RestrepoAún no hay calificaciones
- Monografia Calorimetría AnimalDocumento34 páginasMonografia Calorimetría AnimalEder Aquino Galarza100% (2)
- Signos y Sintomas RespiratoriosDocumento87 páginasSignos y Sintomas RespiratoriosSEBASTIÁN BENÍTEZAún no hay calificaciones
- Guía Signos Vitales y Glosario de Términos Técnicos Profe E.U Chamed S.Documento14 páginasGuía Signos Vitales y Glosario de Términos Técnicos Profe E.U Chamed S.zamar20100% (1)
- Cambios en La AdolescenciaDocumento3 páginasCambios en La AdolescenciaJana Misseth Gaxiola CastroAún no hay calificaciones
- HSSE 6 - Procedimiento Entrega - Revisión EMO y CAMO Rev5 310316Documento15 páginasHSSE 6 - Procedimiento Entrega - Revisión EMO y CAMO Rev5 310316Jimmy David Espinoza MejiaAún no hay calificaciones
- Respiracion y Relajacion en Situaciones de Actividad Fisica o DeportesDocumento3 páginasRespiracion y Relajacion en Situaciones de Actividad Fisica o DeportesLeonardo David Zapardiel RicoAún no hay calificaciones
- Practica Word Belda PDFDocumento10 páginasPractica Word Belda PDFLeonardo BeldaAún no hay calificaciones