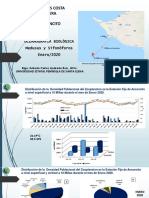
También podría gustarte
- Buelvas Et Al. - Proyecto CampoDocumento10 páginasBuelvas Et Al. - Proyecto CampoDavid Vergara MorenoAún no hay calificaciones
- Texto de Ictiología Lagler PDFDocumento498 páginasTexto de Ictiología Lagler PDFbiologomarino88% (8)
- Concreto EstructuralDocumento38 páginasConcreto EstructuralJohnnyCarbajalGuzman100% (2)
- Articulo CientificoDocumento10 páginasArticulo CientificoLeonteo Te CacheAún no hay calificaciones
- Algas RochaDocumento14 páginasAlgas RochaAle BerniniAún no hay calificaciones
- Diversidad de PlanctonDocumento15 páginasDiversidad de PlanctonAlex RamirezAún no hay calificaciones
- Estructura de La Comunidad de Peces en Cuatro Estuarios Del Pacífico MexicanoDocumento13 páginasEstructura de La Comunidad de Peces en Cuatro Estuarios Del Pacífico MexicanoEdgar SandovalAún no hay calificaciones
- Hidromedusas Del Pacífico Colombiano: Aspectos Básicos de Su Dinámica EcológicaDocumento24 páginasHidromedusas Del Pacífico Colombiano: Aspectos Básicos de Su Dinámica EcológicaEverton GiachiniAún no hay calificaciones
- ReparacionDocumento9 páginasReparacionJuana Ramona GonzálezAún no hay calificaciones
- MacrofitasDocumento12 páginasMacrofitasJuanz Sebastian DiagamaAún no hay calificaciones
- 1483-Texto Del Artículo-8618-2-10-20210201Documento20 páginas1483-Texto Del Artículo-8618-2-10-20210201ESTEFANI KAROLINA DE LA CRUZ ACEVEDOAún no hay calificaciones
- Tea Final ComunidadesDocumento23 páginasTea Final ComunidadesJulia Elena Sánchez LópezAún no hay calificaciones
- CangrejosDocumento19 páginasCangrejosAparoAún no hay calificaciones
- Biodiersidad de Crustáceos Dulceacuicolas Del Centro de Nuevo León y Noroeste de TamaulipasDocumento33 páginasBiodiersidad de Crustáceos Dulceacuicolas Del Centro de Nuevo León y Noroeste de TamaulipasPresa Rodrigo Gómez, "La Boca"Aún no hay calificaciones
- Enfermedades de La PotaDocumento27 páginasEnfermedades de La PotaGuillermo SamuelAún no hay calificaciones
- Taxonomía, Ecologia y Estructura de Las Comunidades de Peces enDocumento439 páginasTaxonomía, Ecologia y Estructura de Las Comunidades de Peces enRicardo Torres CárdenasAún no hay calificaciones
- Art 16Documento15 páginasArt 16MercedesTorresAún no hay calificaciones
- Macroinvertebrados Bentónicos de Dos Lagos de Alta Montaña en El Estado de Morelos, MéxicoDocumento13 páginasMacroinvertebrados Bentónicos de Dos Lagos de Alta Montaña en El Estado de Morelos, MéxicoEULER ROSSEL CAMPOS SALDA�AAún no hay calificaciones
- 10 Epherra ErizodemarArbaciadufresniiyUndariapinnatifidaDocumento12 páginas10 Epherra ErizodemarArbaciadufresniiyUndariapinnatifidaMare Del PacifícoAún no hay calificaciones
- Sterling & Roa - CORREGIDODocumento6 páginasSterling & Roa - CORREGIDOJess Roa VillabonaAún no hay calificaciones
- Zamorano Etal 2006Documento11 páginasZamorano Etal 2006lopezalcuadradoAún no hay calificaciones
- ANCHOVETADocumento46 páginasANCHOVETAfrancojesusAún no hay calificaciones
- Abundancia Dominancia y Diversidad PDFDocumento6 páginasAbundancia Dominancia y Diversidad PDFLunaCharitoAún no hay calificaciones
- Boletin 34 (2) - 9Documento25 páginasBoletin 34 (2) - 9Ricardo Edwin Quispe ZegarraAún no hay calificaciones
- Grupos Recurresntes de Peces y Crustaceos Zona CentralDocumento10 páginasGrupos Recurresntes de Peces y Crustaceos Zona CentralComo MuchoAún no hay calificaciones
- 4274-Texto Del Artículo-14486-1-10-20170610 PDFDocumento38 páginas4274-Texto Del Artículo-14486-1-10-20170610 PDFDaniel Hills SparksAún no hay calificaciones
- ZoonplanctonDocumento14 páginasZoonplanctonStefannyMAún no hay calificaciones
- Herpetos y Manglares, Una Relación Poco ConocidaDocumento7 páginasHerpetos y Manglares, Una Relación Poco ConocidaSara LaniakeaAún no hay calificaciones
- 4415-Texto Del Artículo-16792-1-10-20220429Documento26 páginas4415-Texto Del Artículo-16792-1-10-20220429SaulAún no hay calificaciones
- Primer Registro Del Genero MetadinophysisDocumento16 páginasPrimer Registro Del Genero Metadinophysis6621920001 YOLEINIS ELIZABETH ACOSTA GUERRERO ESTUDIANTE ACTIVOAún no hay calificaciones
- 2013 Los Langostinos Del Género Macrobrachium Con Importancia Económica yDocumento25 páginas2013 Los Langostinos Del Género Macrobrachium Con Importancia Económica yMax MaxAún no hay calificaciones
- Gcfi 64-7Documento5 páginasGcfi 64-7ZakAún no hay calificaciones
- Eulogio SotoDocumento13 páginasEulogio SotoGabrielAún no hay calificaciones
- Revista Investigaciones MarinasDocumento21 páginasRevista Investigaciones MarinasJuan Carlos GarciaAún no hay calificaciones
- 326-347 - Libro - Biodiversidad - Cuba - Capítulo 17Documento22 páginas326-347 - Libro - Biodiversidad - Cuba - Capítulo 17Lucerito Villamil100% (1)
- Patrones de Distribución de Haemulidae A Lo Largo Del Noreste de BrasilDocumento24 páginasPatrones de Distribución de Haemulidae A Lo Largo Del Noreste de Brasilluis medina “el medis”Aún no hay calificaciones
- Laguna El FarallónDocumento4 páginasLaguna El FarallónIván UrangaAún no hay calificaciones
- Estudio Preliminar de Algunos Aspectos Ambientales y Ecologicos de Peces y Macroinvertebrados en El Rio Tutunendo PDFDocumento10 páginasEstudio Preliminar de Algunos Aspectos Ambientales y Ecologicos de Peces y Macroinvertebrados en El Rio Tutunendo PDFGabo MontesAún no hay calificaciones
- Oviedo-Correa L. et al. (2009). Evalucación de los riesgos de afectación por el establecimiento de granjas atuneras en relación con la distribución espacial de cetáceos en el Golfo Dulce, Costa Rica. Rev. Mar.y Cos. 1: 159-174Documento16 páginasOviedo-Correa L. et al. (2009). Evalucación de los riesgos de afectación por el establecimiento de granjas atuneras en relación con la distribución espacial de cetáceos en el Golfo Dulce, Costa Rica. Rev. Mar.y Cos. 1: 159-174Centro de Investigación de Cetáceos (CEIC) - Costa RicaAún no hay calificaciones
- Producción de Hojarasca en El Manglar de La Reserva Estatal de Palmar en SisalDocumento15 páginasProducción de Hojarasca en El Manglar de La Reserva Estatal de Palmar en SisalEnrique Barush Flores FloresAún no hay calificaciones
- Informe FINAL Peces de Manglar Sanquianga - Entrega 16 Jun 14Documento14 páginasInforme FINAL Peces de Manglar Sanquianga - Entrega 16 Jun 14gustavo castellanosAún no hay calificaciones
- Arrecifes Coralinos Isla FuerteDocumento7 páginasArrecifes Coralinos Isla Fuertegustavo AraqueAún no hay calificaciones
- Informe Litoral Rocoso Macroalgas 2023-2Documento18 páginasInforme Litoral Rocoso Macroalgas 2023-2Santiago Alberto Santos NietoAún no hay calificaciones
- Fondos BlandosDocumento4 páginasFondos BlandoslejoumeAún no hay calificaciones
- Tavares 2019 Estado Actual Elasmo Cienc PesqDocumento23 páginasTavares 2019 Estado Actual Elasmo Cienc PesqHelado De 7 semillasAún no hay calificaciones
- Informe 49-1 EvalPatallus2011 - 2019Documento15 páginasInforme 49-1 EvalPatallus2011 - 2019Rafael Alfredo Gonzales BazalarAún no hay calificaciones
- Estructura Poblacional y Estado Reproductivo Del Bentón (Hoplias Malabaricus) en La Laguna de Inundación Urbana "Las Palquitas". Trinidad, Beni, Bolivia 2016.Documento76 páginasEstructura Poblacional y Estado Reproductivo Del Bentón (Hoplias Malabaricus) en La Laguna de Inundación Urbana "Las Palquitas". Trinidad, Beni, Bolivia 2016.Lucas Perez HerediaAún no hay calificaciones
- Diversidad y Cobertura de La Comunidad Bentónica Del Arrecife CA JDocumento5 páginasDiversidad y Cobertura de La Comunidad Bentónica Del Arrecife CA JEstebanAún no hay calificaciones
- Plantemiento Del Problema DescriptivoDocumento4 páginasPlantemiento Del Problema DescriptivoKaren OchoaAún no hay calificaciones
- 78 78 1 PBDocumento13 páginas78 78 1 PBRichard ChavezAún no hay calificaciones
- Biodiversidad MarinaDocumento34 páginasBiodiversidad MarinaMaurilio HuAún no hay calificaciones
- Articulo Abundancia de La Salamandra Ambystoma AndersoniDocumento12 páginasArticulo Abundancia de La Salamandra Ambystoma AndersoniLeonell MorantesAún no hay calificaciones
- Informe Practico de Limnología N°5Documento9 páginasInforme Practico de Limnología N°5Jorhs Garcia MurilloAún no hay calificaciones
- Equinodermos Del Caribe de GuatemalaDocumento8 páginasEquinodermos Del Caribe de GuatemalaSOfi MéndezAún no hay calificaciones
- Equinodermos Del Parque Nacional Isla Isabel, Nayarit, MéxicoDocumento11 páginasEquinodermos Del Parque Nacional Isla Isabel, Nayarit, MéxicoCRISTIAN MOISES GALVAN VILLAAún no hay calificaciones
- Laguna Cfe YalahuaDocumento10 páginasLaguna Cfe Yalahuajose razoAún no hay calificaciones
- Abundancia y Distribución Del Cangrejo Violinista Uca Rapax en La Laguna Las PeoniasDocumento5 páginasAbundancia y Distribución Del Cangrejo Violinista Uca Rapax en La Laguna Las PeoniasSavier F Acosta F100% (1)
- Diversity of The Heteroptera (Insecta:Hemiptera) Aquatic and Semi-Acuatic Assemblage of The Rio Grande Basing (Jujuy-Argentina) in An Altitudinal and Temporal GradientDocumento10 páginasDiversity of The Heteroptera (Insecta:Hemiptera) Aquatic and Semi-Acuatic Assemblage of The Rio Grande Basing (Jujuy-Argentina) in An Altitudinal and Temporal Gradienteugenia contrerasAún no hay calificaciones
- Mantos AlgalesDocumento8 páginasMantos AlgalesGianninaMiguelJuradoAún no hay calificaciones
- 2019 - Pereira Et Al - Variación en Pequeña Escala de Comunidades Macrobentónicas en Laguna CosteraDocumento23 páginas2019 - Pereira Et Al - Variación en Pequeña Escala de Comunidades Macrobentónicas en Laguna CosteraPablo NayaAún no hay calificaciones
- Ecología y conservación en los mares de MéxicoDe EverandEcología y conservación en los mares de MéxicoAún no hay calificaciones
- Programas de Protección Regional de Áreas Prioritarias de grandes ballenas en el golfo de California y costa occidental de Baja California Sur: Propuestas de conservaciónDe EverandProgramas de Protección Regional de Áreas Prioritarias de grandes ballenas en el golfo de California y costa occidental de Baja California Sur: Propuestas de conservaciónAún no hay calificaciones
- Quetognatos Del Golfo de GuayaquilDocumento60 páginasQuetognatos Del Golfo de GuayaquilbiologomarinoAún no hay calificaciones
- Boletín Erfen 31 Enero 2020Documento6 páginasBoletín Erfen 31 Enero 2020biologomarinoAún no hay calificaciones
- Indicadores Biológicos-Upse Enero 2020Documento7 páginasIndicadores Biológicos-Upse Enero 2020biologomarinoAún no hay calificaciones
- Estractificación Vertical Del OceanoDocumento3 páginasEstractificación Vertical Del OceanobiologomarinoAún no hay calificaciones
- Boletín Del Comité Erfen 2018Documento5 páginasBoletín Del Comité Erfen 2018biologomarinoAún no hay calificaciones
- 2014 Lavaniegos BGNF-enprensa PDFDocumento27 páginas2014 Lavaniegos BGNF-enprensa PDFbiologomarinoAún no hay calificaciones
- 2014 Lavaniegos BGNF-enprensa PDFDocumento27 páginas2014 Lavaniegos BGNF-enprensa PDFbiologomarinoAún no hay calificaciones
- DC1400 IM Spanish-122011Documento85 páginasDC1400 IM Spanish-122011biologomarinoAún no hay calificaciones
- 2014 Lavaniegos BGNF-enprensa PDFDocumento27 páginas2014 Lavaniegos BGNF-enprensa PDFbiologomarinoAún no hay calificaciones
- 2014 Lavaniegos BGNF-enprensa PDFDocumento27 páginas2014 Lavaniegos BGNF-enprensa PDFbiologomarinoAún no hay calificaciones
- ScieloDocumento17 páginasScieloLAZYAún no hay calificaciones
- Clase 4 SifonòforosDocumento29 páginasClase 4 SifonòforosbiologomarinoAún no hay calificaciones
- Ejercicios CuencaDocumento10 páginasEjercicios CuencaKiara Neyra SotoAún no hay calificaciones
- AfloramientosDocumento20 páginasAfloramientosJose Armando Eccoña TelloAún no hay calificaciones
- Revista - 81 MYSIDACEOSDocumento10 páginasRevista - 81 MYSIDACEOSbiologomarinoAún no hay calificaciones
- Capitulo4 Determinacion Del EscurrimientoDocumento14 páginasCapitulo4 Determinacion Del EscurrimientoRaul Hernandez LopezAún no hay calificaciones
- Atlas Org Planctonicos 1Documento253 páginasAtlas Org Planctonicos 1Marce Lino Lopez Rendon100% (3)
- Cuenca SDocumento30 páginasCuenca SrutAún no hay calificaciones
- Clave Ilustrada Del Género MysidaceoDocumento21 páginasClave Ilustrada Del Género MysidaceobiologomarinoAún no hay calificaciones
- Catalogo de Fitoplancton Cartagena2012Documento136 páginasCatalogo de Fitoplancton Cartagena2012Naza RioAún no hay calificaciones
- Ictiologia General MadrigalDocumento7 páginasIctiologia General MadrigalbiologomarinoAún no hay calificaciones
- Practica 5 IctiologiaDocumento2 páginasPractica 5 IctiologiabiologomarinoAún no hay calificaciones
- Act. Práctica # 2Documento2 páginasAct. Práctica # 2Sara Isabel Robleto ValdiviaAún no hay calificaciones
- Semejanza de TurbomáquinasDocumento18 páginasSemejanza de TurbomáquinasRogerAún no hay calificaciones
- Wilman Leonel Ureña Castro - Asignacion de La Semana Ii-CartografiaDocumento9 páginasWilman Leonel Ureña Castro - Asignacion de La Semana Ii-CartografiaWilman UreñaAún no hay calificaciones
- Boris Laboratorio 7777Documento14 páginasBoris Laboratorio 7777CEIAQP EIRLAún no hay calificaciones
- Importancia de La Seguridad en Resonancia Magnética: Jaime Torres Juárez, Silvia Hidalgo TobónDocumento14 páginasImportancia de La Seguridad en Resonancia Magnética: Jaime Torres Juárez, Silvia Hidalgo TobónLucía MoraAún no hay calificaciones
- Operacionalización de Variables: Odontología Activa Revista Científica January 2019Documento7 páginasOperacionalización de Variables: Odontología Activa Revista Científica January 2019Jorge SanchezAún no hay calificaciones
- Trabajo Practico N 2 de Algebra Lineal 2Documento11 páginasTrabajo Practico N 2 de Algebra Lineal 2Francisco PolzoniAún no hay calificaciones
- Práctica 2-Lab Mec MaterialesDocumento4 páginasPráctica 2-Lab Mec MaterialesCristopher CalocaAún no hay calificaciones
- Marco Montalvo - José Meléndez PalmaDocumento29 páginasMarco Montalvo - José Meléndez PalmaGunder Huacho VaraAún no hay calificaciones
- Informe - 2 - ELECTROSTÁTICADocumento11 páginasInforme - 2 - ELECTROSTÁTICAGeraldin Ching FrancoAún no hay calificaciones
- 06 Subconsultas y Operadores de ConjuntosDocumento11 páginas06 Subconsultas y Operadores de ConjuntosCristhian DiazAún no hay calificaciones
- Ciencias Nat - Guia 2 - 10 y 11 ListaDocumento8 páginasCiencias Nat - Guia 2 - 10 y 11 Listaddanovis0629Aún no hay calificaciones
- Proteínas de Interés Biológico BB 2410Documento1 páginaProteínas de Interés Biológico BB 2410Sofia Beltran RuizAún no hay calificaciones
- NB6 Prueba Lenguaje Algebraico y Ecuaciones BDocumento3 páginasNB6 Prueba Lenguaje Algebraico y Ecuaciones BNorma Elena RojasAún no hay calificaciones
- Características Del Agua de CalderasDocumento7 páginasCaracterísticas Del Agua de CalderasJean CarLosAún no hay calificaciones
- Formacion de La OrinaDocumento5 páginasFormacion de La OrinaFernanda OvandoAún no hay calificaciones
- Actividad 3 Calculo Diferencial e Integral ResueltoDocumento11 páginasActividad 3 Calculo Diferencial e Integral Resueltoedgar jafet murillo silvaAún no hay calificaciones
- Examen 9no.Documento6 páginasExamen 9no.KELVIN DIAZAún no hay calificaciones
- Universidad Nacional Daniel Alcides Carrión 2018Documento5 páginasUniversidad Nacional Daniel Alcides Carrión 2018Jhon Hrnrry Reyes HuamanAún no hay calificaciones
- Guía Práctica para Diseño de Encofrados Verticales de MaderaDocumento9 páginasGuía Práctica para Diseño de Encofrados Verticales de MaderaJohn MejiaAún no hay calificaciones
- Tema 4 - Productos Notables - 10!04!2018Documento4 páginasTema 4 - Productos Notables - 10!04!2018Erick Perez Castillo100% (1)
- Deformación Por CorteDocumento19 páginasDeformación Por Cortepedro valenciaAún no hay calificaciones
- Probabilístico, Mapa Mental!Documento2 páginasProbabilístico, Mapa Mental!Wendy ChiAún no hay calificaciones
- Actividades-Semana 4-1Documento3 páginasActividades-Semana 4-1sulannee alejoAún no hay calificaciones
- TALLER TEOREMA DE LAPLACE Y BAYES SoloDocumento13 páginasTALLER TEOREMA DE LAPLACE Y BAYES SoloDayana FuentesAún no hay calificaciones
- LAB - Ospf - Clase2 - Jennyfer HernandezDocumento3 páginasLAB - Ospf - Clase2 - Jennyfer HernandezJennyfer HernandezAún no hay calificaciones
- TP Laboratorio QuimicaDocumento5 páginasTP Laboratorio QuimicaJavier R Hernández100% (2)
- Resumen Estadística 3Documento102 páginasResumen Estadística 3Tobias LuceroAún no hay calificaciones
- LACCEI 2020 - Gonzalez - Aguilar - Huaco PDFDocumento6 páginasLACCEI 2020 - Gonzalez - Aguilar - Huaco PDFElio VidalAún no hay calificaciones