También podría gustarte
- Los discos. Adquisición, cría, últimas variedades seleccionadas, prevención y cura de las enfermedadesDe EverandLos discos. Adquisición, cría, últimas variedades seleccionadas, prevención y cura de las enfermedadesCalificación: 4.5 de 5 estrellas4.5/5 (2)
- CATALOGO MERLUZAS 4ta ParteDocumento9 páginasCATALOGO MERLUZAS 4ta ParteMartin RequeAún no hay calificaciones
- Mamíferos en peligro. Entre ballenas, delfines, zifios y manatíesDe EverandMamíferos en peligro. Entre ballenas, delfines, zifios y manatíesAún no hay calificaciones
- CATALOGO MERLUZAS 2da ParteDocumento9 páginasCATALOGO MERLUZAS 2da ParteMartin RequeAún no hay calificaciones
- Clave para La Identificacion de Los Calliphoridae de La Républica Argentina (Diptera)Documento4 páginasClave para La Identificacion de Los Calliphoridae de La Républica Argentina (Diptera)Flor Galván SánchezAún no hay calificaciones
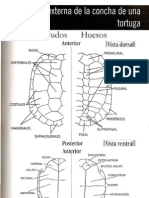
- Clave TestudinesDocumento2 páginasClave TestudinesJavier Aldo BianchiAún no hay calificaciones
- Clave Gusano BlancoDocumento5 páginasClave Gusano BlancoJuan Carlos M CAún no hay calificaciones
- Biologia PesqueraDocumento18 páginasBiologia PesqueraJDGCAún no hay calificaciones
- 1.2 10 Guia - IdentificacionDocumento65 páginas1.2 10 Guia - IdentificacionDavid Esteban Davila YepesAún no hay calificaciones
- Caso 1978Documento16 páginasCaso 1978Franvier Val-SanAún no hay calificaciones
- Guia de Peces Catilaginosos en El Río de La Plata y Su Frente Oceánico PDFDocumento61 páginasGuia de Peces Catilaginosos en El Río de La Plata y Su Frente Oceánico PDFAye IñigoAún no hay calificaciones
- Clave Larvas Tricoptero PDFDocumento8 páginasClave Larvas Tricoptero PDFClaudia SanchezAún no hay calificaciones
- Guía Ilustrada de Peces de Isla Del Coco y Arrecifes de La Costa Pacífica de América CentralDocumento161 páginasGuía Ilustrada de Peces de Isla Del Coco y Arrecifes de La Costa Pacífica de América CentralTribu GlobalAún no hay calificaciones
- 3 Kath TiburonesDocumento5 páginas3 Kath TiburonesKatherin RendonAún no hay calificaciones
- Claves para La Identificación de Los Peces de Agua DulceDocumento3 páginasClaves para La Identificación de Los Peces de Agua DulcelabraxaslAún no hay calificaciones
- Guia de CrustáceosDocumento8 páginasGuia de CrustáceosDavid F. RiosAún no hay calificaciones
- Tareas Del ViernesDocumento5 páginasTareas Del Viernesmichellearreaga9Aún no hay calificaciones
- Claves Tortugas Marinas, Terrestres & DulceacuicolasDocumento15 páginasClaves Tortugas Marinas, Terrestres & DulceacuicolasCristiam Albeiro FernandezAún no hay calificaciones
- Claves Lombrices de TierraDocumento7 páginasClaves Lombrices de TierraLaura Daniela Sierra GuevaraAún no hay calificaciones
- Claves de KinosternonDocumento3 páginasClaves de KinosternonFrancisco CabreraAún no hay calificaciones
- AGUJADocumento56 páginasAGUJAtalaradeiAún no hay calificaciones
- Cuadernillopracticas 2Documento39 páginasCuadernillopracticas 2oooMaisi LeonoooAún no hay calificaciones
- Tareas Del ViernesDocumento5 páginasTareas Del Viernesmichellearreaga9Aún no hay calificaciones
- TP 9 Clave Lepidosauria 2015Documento17 páginasTP 9 Clave Lepidosauria 2015nenriquezaAún no hay calificaciones
- Tema 10.3Documento42 páginasTema 10.3Manolo Dominguez BecerraAún no hay calificaciones
- ReduviidaeDocumento17 páginasReduviidaeArturo LopezAún no hay calificaciones
- 10 CrustaceaDocumento40 páginas10 CrustaceaPaula MadridAún no hay calificaciones
- Claves Peces Agua Dulce 1Documento134 páginasClaves Peces Agua Dulce 1AnGy Katy MarsAún no hay calificaciones
- Los Peces Oseos, Claves Dicotómicas y Peces Comunes en El PerúDocumento8 páginasLos Peces Oseos, Claves Dicotómicas y Peces Comunes en El PerúRomulo Aycachi Inga89% (9)
- Jaibas PDFDocumento30 páginasJaibas PDFOscar Lopez CubillosAún no hay calificaciones
- Clave de Arañas Gerschman SchiapelliDocumento30 páginasClave de Arañas Gerschman SchiapelliBOCAún no hay calificaciones
- Roman-Valencia 2005 Bryconamericus Venezuela & EcuadorDocumento26 páginasRoman-Valencia 2005 Bryconamericus Venezuela & EcuadorcarlochessAún no hay calificaciones
- Inverte CompiladoDocumento57 páginasInverte CompiladoShizui LanAún no hay calificaciones
- Libro ChaetognathaDocumento94 páginasLibro Chaetognathafabiola villa alvaradoAún no hay calificaciones
- Práctica 8 CrustaceaDocumento22 páginasPráctica 8 CrustaceaLaura APAún no hay calificaciones
- AnfibiosDocumento39 páginasAnfibios34rtghu765reAún no hay calificaciones
- Anfibios PDFDocumento39 páginasAnfibios PDFHamilton Alexi Tuesta DazaAún no hay calificaciones
- BacoretaDocumento8 páginasBacoretafcocajaAún no hay calificaciones
- Aspectos Recolección, de MoluscosDocumento18 páginasAspectos Recolección, de MoluscosarrozpolloAún no hay calificaciones
- Caso 2 - Maybe - FerroDocumento8 páginasCaso 2 - Maybe - FerromaytumbleAún no hay calificaciones
- Práctica #02 Regiones Del Cuerpo de Los PecesDocumento6 páginasPráctica #02 Regiones Del Cuerpo de Los Peces21031b0561Aún no hay calificaciones
- Parasitoides de Mosca PrietaDocumento8 páginasParasitoides de Mosca PrietaOliverBarrientosSidaAún no hay calificaciones
- 5 - RRHH Moluscos 2020-1Documento85 páginas5 - RRHH Moluscos 2020-1briggitte aquinoAún no hay calificaciones
- Clave de CrustaceosDocumento9 páginasClave de Crustaceosatosky15Aún no hay calificaciones
- Invertebrados Marinos de La Región La LibertadDocumento110 páginasInvertebrados Marinos de La Región La LibertadSarita Virginia Campos LeónAún no hay calificaciones
- 0.C Coleoptera BDocumento7 páginas0.C Coleoptera Braitomi04Aún no hay calificaciones
- Peces de Nicaragua - 03Documento29 páginasPeces de Nicaragua - 03Danfer Jossiel Duarte BelloAún no hay calificaciones
- 5 - RRHH Peces Generalidades 2022-1Documento69 páginas5 - RRHH Peces Generalidades 2022-1Diego Enrique Mamani CubaAún no hay calificaciones
- PALANGREDocumento18 páginasPALANGRERAMOS RUBIO DIEGO ANDERSONAún no hay calificaciones
- Caracteres Meristicos ActualizadoDocumento30 páginasCaracteres Meristicos Actualizadolauvamal9860% (5)
- Nociones Basicas de Los PecesDocumento14 páginasNociones Basicas de Los Pecessafer1411Aún no hay calificaciones
- Album CamaronesDocumento52 páginasAlbum CamaronesSharon Abigail UrbinaAún no hay calificaciones
- PHYLUM ARTHTOPODA PARTE 4. MaxilopodosDocumento18 páginasPHYLUM ARTHTOPODA PARTE 4. MaxilopodosChristian Cabrera VidalAún no hay calificaciones
- Familia Cichlidae DFDocumento66 páginasFamilia Cichlidae DFLianka GarciaAún no hay calificaciones
- LonchaeidaeDocumento49 páginasLonchaeidaerodrigosotoandradesAún no hay calificaciones
- Saurios Colombia ISSUUDocumento80 páginasSaurios Colombia ISSUUjose david sanchez salgadoAún no hay calificaciones
- Morfologia Externa e Interna de Panalirus Gracilis Querevalu Dioses Jose Luis 2017Documento21 páginasMorfologia Externa e Interna de Panalirus Gracilis Querevalu Dioses Jose Luis 2017jose suncion50% (2)
- CONICET Digital Nro. ADocumento15 páginasCONICET Digital Nro. ALuana AvendañoAún no hay calificaciones
- Revista El Hornero, Volumen 2, #4. 1922.Documento91 páginasRevista El Hornero, Volumen 2, #4. 1922.Fernando Julio BioléAún no hay calificaciones
- Amphibia - Rodriguez, Andrea A.Documento11 páginasAmphibia - Rodriguez, Andrea A.Lucas VillagranAún no hay calificaciones
- TESISDocumento70 páginasTESISMartin RequeAún no hay calificaciones
- Elementos de Importancia EconómicaDocumento3 páginasElementos de Importancia EconómicaMartin RequeAún no hay calificaciones
- Massa 2006 PDFDocumento236 páginasMassa 2006 PDFpikipoliAún no hay calificaciones
- Catalogo MerluzasDocumento12 páginasCatalogo MerluzasMartin RequeAún no hay calificaciones
- CATALOGO MERLUZAS 5ta ParteDocumento9 páginasCATALOGO MERLUZAS 5ta ParteMartin RequeAún no hay calificaciones
- Microorganismos MarcadoresDocumento20 páginasMicroorganismos MarcadoresJohn Fernando Galvez100% (1)
- Decreto825338-2009 - Reglamento Ley26427 PDFDocumento9 páginasDecreto825338-2009 - Reglamento Ley26427 PDFMartin RequeAún no hay calificaciones
- Decreto-1374 Pasantias NACIÓN PDFDocumento11 páginasDecreto-1374 Pasantias NACIÓN PDFMartin RequeAún no hay calificaciones
- Ley 26427 Nacion PasantiasDocumento3 páginasLey 26427 Nacion PasantiasMartin RequeAún no hay calificaciones
- Res. 3779 HyS PDFDocumento28 páginasRes. 3779 HyS PDFMartin RequeAún no hay calificaciones
- Hongos de Enfermedades de Postcosecha en Frutos de Cucurbitáceas de Interés Económico de Zonas de Distribución de Trujillo, Perú.Documento6 páginasHongos de Enfermedades de Postcosecha en Frutos de Cucurbitáceas de Interés Económico de Zonas de Distribución de Trujillo, Perú.Martin RequeAún no hay calificaciones
- Olimp Adas Reglamento 2018 PDFDocumento19 páginasOlimp Adas Reglamento 2018 PDFMartin RequeAún no hay calificaciones
- Control de Plagas Industria AlimentariaDocumento18 páginasControl de Plagas Industria AlimentariaAnyi Daniela100% (2)
- Ejercicios de QuímicaDocumento3 páginasEjercicios de QuímicaTammy RodgersAún no hay calificaciones
- VDocumento10 páginasVWilo JaraAún no hay calificaciones
- 2 DaviniDocumento9 páginas2 DaviniMartin RequeAún no hay calificaciones
- Manual en Español SPD SDocumento138 páginasManual en Español SPD Spercydrums50% (2)
- Manual de Estrategias de Enseñanza Aprendizaje - SENA - Cap 1 BisDocumento3 páginasManual de Estrategias de Enseñanza Aprendizaje - SENA - Cap 1 BisMartin RequeAún no hay calificaciones
- Nomenclador de CarrerasDocumento5 páginasNomenclador de CarrerasMartin RequeAún no hay calificaciones
- PolarimetriaDocumento13 páginasPolarimetriaMartin Reque100% (4)
- Acto Dia de La Musica (Nacho)Documento10 páginasActo Dia de La Musica (Nacho)Rosario Moreno60% (5)
- Cuantificacion de Vitamina CDocumento5 páginasCuantificacion de Vitamina CAbraham Morales100% (1)
- Decreto 4955Documento6 páginasDecreto 4955Martin RequeAún no hay calificaciones
- Ley 26427 Nacion PasantiasDocumento3 páginasLey 26427 Nacion PasantiasMartin RequeAún no hay calificaciones
- Decreto 4955Documento6 páginasDecreto 4955Martin RequeAún no hay calificaciones
- Microorganismos MarcadoresDocumento20 páginasMicroorganismos MarcadoresJohn Fernando Galvez100% (1)
- Tesis Sobre SalchichasDocumento401 páginasTesis Sobre SalchichasMartin RequeAún no hay calificaciones
- Tema 2-Músculos Del Hombro - 2020-2021 AVDocumento22 páginasTema 2-Músculos Del Hombro - 2020-2021 AVOscar Espinosa GarciaAún no hay calificaciones
- CUENTOSDocumento3 páginasCUENTOSRaul de Jesus Montes GonzalezAún no hay calificaciones
- TripticoDocumento4 páginasTripticoHely SocorroAún no hay calificaciones
- Aves Del Uruguay 7Documento18 páginasAves Del Uruguay 7CAroliniAún no hay calificaciones
- Peces GruppyDocumento7 páginasPeces GruppyPatricia Guirao SalcedoAún no hay calificaciones
- Memorama de Animales MinDocumento22 páginasMemorama de Animales MinIvonne RuvalcabaAún no hay calificaciones
- Artropodos y GeneralidadesDocumento7 páginasArtropodos y GeneralidadesMariana Quiñonez BonillaAún no hay calificaciones
- Zoologia 2Documento8 páginasZoologia 2Qjfoñ 0183prearsAún no hay calificaciones
- Carlos La HormigaDocumento4 páginasCarlos La HormigaFrancisco JiménezAún no hay calificaciones
- Listamayoreo 44Documento4 páginasListamayoreo 44Reimundo FuentesAún no hay calificaciones
- Musculos Del CuelloDocumento47 páginasMusculos Del CuelloVictor RiveraAún no hay calificaciones
- Accidentes OseosDocumento3 páginasAccidentes OseosEljuan CalderonAún no hay calificaciones
- 2°claseDocumento78 páginas2°claseAbraham SantuchoAún no hay calificaciones
- AjoloteDocumento13 páginasAjoloteAlejandro Flores Velasco100% (1)
- Anatomía Del Hueso Del Húmero Del Perro AnatomyLearner - El Lugar para Aprender Anatomía Veterinaria en LíneaDocumento36 páginasAnatomía Del Hueso Del Húmero Del Perro AnatomyLearner - El Lugar para Aprender Anatomía Veterinaria en Líneaclips and chipsAún no hay calificaciones
- Fauna Quinta RegiónDocumento17 páginasFauna Quinta Regiónjavdbd havbfoaAún no hay calificaciones
- Tabla para Claves TaxonomicaDocumento2 páginasTabla para Claves TaxonomicaDIEGO SCHMALBACHAún no hay calificaciones
- Exploración FísicaDocumento10 páginasExploración FísicaÅreanną De los ÁngelesAún no hay calificaciones
- Cuento Corto InfantilDocumento2 páginasCuento Corto Infantilgiselle rodriguezAún no hay calificaciones
- Expocicion de BiologiaDocumento12 páginasExpocicion de BiologiaJosue Ramirez MorenoAún no hay calificaciones
- Músculo Pterigoideo Medial HLDocumento7 páginasMúsculo Pterigoideo Medial HLLoren SolisAún no hay calificaciones
- 3Documento9 páginas3Boris VacaAún no hay calificaciones
- Patas Largas y Patas CortasDocumento6 páginasPatas Largas y Patas CortasMelycha EchevarriaAún no hay calificaciones
- Morfologia Pulpar en TemporalesDocumento4 páginasMorfologia Pulpar en TemporalesBLANCAAún no hay calificaciones
- HEMIMETABOLOS Ephemeroptera Odonata PlecopteraDocumento16 páginasHEMIMETABOLOS Ephemeroptera Odonata PlecopteraRamsam JoanfranAún no hay calificaciones
- El Que No Corre VuelaDocumento12 páginasEl Que No Corre VuelaCam AzparrenAún no hay calificaciones
- Animalia - Wikipedia, La Enciclopedia LibreDocumento12 páginasAnimalia - Wikipedia, La Enciclopedia LibreGabriela Silfa GomezAún no hay calificaciones
- Labo 3 y 4 Platelmintos NemátodosDocumento3 páginasLabo 3 y 4 Platelmintos NemátodosVanna VerarutAún no hay calificaciones
- Infografia: AbadejoDocumento1 páginaInfografia: AbadejoMariano VortexAún no hay calificaciones
- Subphylum ChelicerataDocumento74 páginasSubphylum ChelicerataPIERO ALESSANDRO ALFEREZ ABARCAAún no hay calificaciones
- Tropical Reef Fishes: Periplus Nature GuideDe EverandTropical Reef Fishes: Periplus Nature GuideCalificación: 3 de 5 estrellas3/5 (1)
- Introduction to the Practice of Fishery Science, Revised Edition: Revised EditionDe EverandIntroduction to the Practice of Fishery Science, Revised Edition: Revised EditionAún no hay calificaciones
- The Arctic Cod: A Study of Research Into the British Trawl FisheriesDe EverandThe Arctic Cod: A Study of Research Into the British Trawl FisheriesAún no hay calificaciones
- Guidelines for the Fisheries and Aquaculture Sector on Damage and Needs Assessments in EmergenciesDe EverandGuidelines for the Fisheries and Aquaculture Sector on Damage and Needs Assessments in EmergenciesAún no hay calificaciones
- Handbook of Fish Biology and Fisheries: Fish BiologyDe EverandHandbook of Fish Biology and Fisheries: Fish BiologyAún no hay calificaciones
- Understanding Life Below the Beautiful Sea | Children's Fish & Marine LifeDe EverandUnderstanding Life Below the Beautiful Sea | Children's Fish & Marine LifeAún no hay calificaciones
- Fishes and Forestry: Worldwide Watershed Interactions and ManagementDe EverandFishes and Forestry: Worldwide Watershed Interactions and ManagementThomas G. NorthcoteAún no hay calificaciones
- Fishes of the Last Frontier: Life Histories, Biology, Ecology, and Management of Alaska's FishesDe EverandFishes of the Last Frontier: Life Histories, Biology, Ecology, and Management of Alaska's FishesAún no hay calificaciones
- Catch Documentation Schemes for Deep-Sea Fisheries in the ABNJ: Their Value, and Options for Implementation: FAO Fisheries and Aquaculture Technical Paper No. 629De EverandCatch Documentation Schemes for Deep-Sea Fisheries in the ABNJ: Their Value, and Options for Implementation: FAO Fisheries and Aquaculture Technical Paper No. 629Aún no hay calificaciones
- The State of World Fisheries and Aquaculture (Sofia)De EverandThe State of World Fisheries and Aquaculture (Sofia)Aún no hay calificaciones
- Aquaculture VirologyDe EverandAquaculture VirologyFrederick S.B. KibengeAún no hay calificaciones
- Vanishing Fish: Shifting Baselines and the Future of Global FisheriesDe EverandVanishing Fish: Shifting Baselines and the Future of Global FisheriesCalificación: 4.5 de 5 estrellas4.5/5 (2)
- Fishing Operations. 3. Best Practices to Improve Safety at Sea in the Fisheries SectorDe EverandFishing Operations. 3. Best Practices to Improve Safety at Sea in the Fisheries SectorCalificación: 4 de 5 estrellas4/5 (1)
- The New Fish Wave: How to Ignite the Seafood IndustryDe EverandThe New Fish Wave: How to Ignite the Seafood IndustryAún no hay calificaciones
- Fish and Wildlife Management: A Handbook for Mississippi LandownersDe EverandFish and Wildlife Management: A Handbook for Mississippi LandownersCalificación: 5 de 5 estrellas5/5 (1)
- Captain Phil Harris: The Legendary Crab Fisherman, Our Hero, Our DadDe EverandCaptain Phil Harris: The Legendary Crab Fisherman, Our Hero, Our DadCalificación: 3.5 de 5 estrellas3.5/5 (2)
- Saltwater Aquarium Models: Recipes for Creating Beautiful Aquariums That ThriveDe EverandSaltwater Aquarium Models: Recipes for Creating Beautiful Aquariums That ThriveAún no hay calificaciones
- Fishes: Animal Group Science Book For Kids | Children's Zoology Books EditionDe EverandFishes: Animal Group Science Book For Kids | Children's Zoology Books EditionAún no hay calificaciones
- City FishingDe EverandCity FishingJudith SchnellCalificación: 5 de 5 estrellas5/5 (2)