También podría gustarte
- Ecografía normal del árbol urinario y genitales externosDe EverandEcografía normal del árbol urinario y genitales externosAún no hay calificaciones
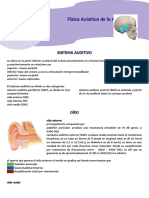
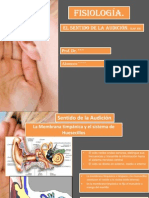
- Capítulo 53 Sentido de La AudiciónDocumento12 páginasCapítulo 53 Sentido de La AudiciónAlonso LunaAún no hay calificaciones
- Fisiología de La Audición.Documento67 páginasFisiología de La Audición.Cecilia Vitale SalasAún no hay calificaciones
- Resumen Sentido de La Audicion Cap. 53Documento5 páginasResumen Sentido de La Audicion Cap. 53nena mishAún no hay calificaciones
- Fisiologia de AudioDocumento3 páginasFisiologia de AudioVerdi ServiciosAún no hay calificaciones
- Fisiología de la audición: transmisión y procesamiento del sonidoDocumento62 páginasFisiología de la audición: transmisión y procesamiento del sonidoCaroline RibeiroAún no hay calificaciones
- Sistema Auditivo PeriféricoDocumento11 páginasSistema Auditivo PeriféricoFabián RuizAún no hay calificaciones
- Fisiologia AudiologiaDocumento62 páginasFisiologia Audiologia10969403Aún no hay calificaciones
- Fisiologia de La Audicion en El Oido ExternoDocumento4 páginasFisiologia de La Audicion en El Oido ExternoESTEFANIA JIMENEZ RINCONESAún no hay calificaciones
- PetroleoDocumento4 páginasPetroleoMaikol Joao De Abreu De Abreu 2do Año AAún no hay calificaciones
- El Sentido de La AudiciónDocumento13 páginasEl Sentido de La AudicióngabrielaAún no hay calificaciones
- Sistema auditivo: apuntes de neurofisiologíaDocumento9 páginasSistema auditivo: apuntes de neurofisiologíaJuan Pablo MoyaAún no hay calificaciones
- Fisiologia AuditivaDocumento8 páginasFisiologia AuditivaSebastián Escobedo100% (1)
- Conducción del sonido desde la membrana timpánica hasta la cócleaDocumento6 páginasConducción del sonido desde la membrana timpánica hasta la cócleaSarely FernándezAún no hay calificaciones
- 2. HIPOACUSIA.Documento14 páginas2. HIPOACUSIA.Diana BenitezAún no hay calificaciones
- Función de la trompa de EustaquioDocumento19 páginasFunción de la trompa de EustaquioDavid GutiérrezAún no hay calificaciones
- 4 Fisiología Auditiva PDFDocumento62 páginas4 Fisiología Auditiva PDFflga. katherine aguirreAún no hay calificaciones
- Guía Final FisiologíaDocumento32 páginasGuía Final FisiologíaAriana Francella Mercado LópezAún no hay calificaciones
- Biofisica de La Audicion 2015Documento69 páginasBiofisica de La Audicion 2015Roosvelt Cruz CórdovaAún no hay calificaciones
- Estructuras Del OídoDocumento3 páginasEstructuras Del OídoMia OchoaAún no hay calificaciones
- Física Acustica de La AudiciónDocumento7 páginasFísica Acustica de La Audicióndenisse.carvalhoAún no hay calificaciones
- Resumen OidoDocumento5 páginasResumen OidoSilvina GrassoAún no hay calificaciones
- Biofísica: Surge después de las radiaciones nuclearesDocumento16 páginasBiofísica: Surge después de las radiaciones nuclearesJesafAún no hay calificaciones
- Fisiologia - Audicion Cap. 52Documento24 páginasFisiologia - Audicion Cap. 52Wander Romero Martins67% (3)
- Audición: Transmisión del sonido a través del oído medioDocumento4 páginasAudición: Transmisión del sonido a través del oído medioJuliette RamírezAún no hay calificaciones
- 3 - El Hombre y El Ruido - 220526 - 190242Documento13 páginas3 - El Hombre y El Ruido - 220526 - 190242Oscar GarzonAún no hay calificaciones
- Cuestinario para El Examen Final - Parte IDocumento14 páginasCuestinario para El Examen Final - Parte IJomarisa VargasAún no hay calificaciones
- El Mec. de La Percepción y Sus Caract. Cap 2Documento110 páginasEl Mec. de La Percepción y Sus Caract. Cap 2nano261191Aún no hay calificaciones
- Sistema auditivo: introducción a las propiedades de las ondas sonoras y la audiciónDocumento6 páginasSistema auditivo: introducción a las propiedades de las ondas sonoras y la audiciónDenis Peralta VillalobosAún no hay calificaciones
- Trab Neuro1Documento9 páginasTrab Neuro1Jessica RiveraAún no hay calificaciones
- Biofisica AudicionDocumento15 páginasBiofisica AudicionAlessandra AsencioAún no hay calificaciones
- Resumen HigieneDocumento20 páginasResumen HigieneMatias GomezAún no hay calificaciones
- BIOFISICA DE LA AUDICIONDocumento8 páginasBIOFISICA DE LA AUDICIONCATALINA NANIAún no hay calificaciones
- Fisiología del oído medioDocumento6 páginasFisiología del oído medioDiego Chavez ZumaetaAún no hay calificaciones
- Anatomía y fisiología del oídoDocumento22 páginasAnatomía y fisiología del oídoBruno OsorioAún no hay calificaciones
- Sonido y La Audicion BiofisicaDocumento7 páginasSonido y La Audicion BiofisicaBrittany ContreraAún no hay calificaciones
- Seminario 13 Fisica Del SonidoDocumento6 páginasSeminario 13 Fisica Del SonidoAnonymous m43RBxOVZ2Aún no hay calificaciones
- Fisica de La AudicionDocumento5 páginasFisica de La AudicionKarlaPatriciaAún no hay calificaciones
- Sistema Sensorial de La Audición 2Documento35 páginasSistema Sensorial de La Audición 2Fernando Junior Parra UchasaraAún no hay calificaciones
- Audición - HoussayDocumento20 páginasAudición - HoussayJorgeRodriguezAún no hay calificaciones
- Fisiología de la audición y el vestíbuloDocumento37 páginasFisiología de la audición y el vestíbuloHugo OrtegaAún no hay calificaciones
- Tratado de Medicina Guyton Ty Hall-Cap 52 ResumenDocumento18 páginasTratado de Medicina Guyton Ty Hall-Cap 52 ResumenLudwing Pérez100% (2)
- Cuadro Comparativo OidoDocumento4 páginasCuadro Comparativo OidoMaggy HernandezAún no hay calificaciones
- Tema 8 FADocumento6 páginasTema 8 FA心夏老奶Aún no hay calificaciones
- Tema 2 EL MECANISMO DE LA AUDICIÓNDocumento10 páginasTema 2 EL MECANISMO DE LA AUDICIÓNx3mgzAún no hay calificaciones
- Sistema AuditivoDocumento11 páginasSistema Auditivomelanie palaciosAún no hay calificaciones
- Biofisica P3 S.14Documento2 páginasBiofisica P3 S.14Denilson MendozaAún no hay calificaciones
- Ruido y vibraciones: La naturaleza del sonido y la audición humanaDocumento46 páginasRuido y vibraciones: La naturaleza del sonido y la audición humanaLiz AnnaAún no hay calificaciones
- Audición - PurvesDocumento18 páginasAudición - PurvesJochiJaramilloAún no hay calificaciones
- Fisiología AuditivaDocumento14 páginasFisiología AuditivaVivi SeguraAún no hay calificaciones
- Percepción de Los SonidosDocumento4 páginasPercepción de Los SonidosDarvin MateoAún no hay calificaciones
- Biofisica de La AudicionDocumento74 páginasBiofisica de La AudicionJafreisy Abigail Reynoso MoyaAún no hay calificaciones
- TP Higiene 4 LucasDocumento4 páginasTP Higiene 4 LucasMatias GomezAún no hay calificaciones
- Fisiología de La AudiciónDocumento13 páginasFisiología de La AudicióncamilaAún no hay calificaciones
- AudiometriaDocumento24 páginasAudiometriaMary Nicole Aráuz AráuzAún no hay calificaciones
- Anatomia y Fisiologia de La AudicionDocumento5 páginasAnatomia y Fisiologia de La AudicionJaume Lazaro Hernandez100% (1)
- Fisiología Del OídoDocumento30 páginasFisiología Del OídoMaggieShulzAún no hay calificaciones
- El Sentido de La AudiciónDocumento4 páginasEl Sentido de La AudiciónJeffrey LoayzaAún no hay calificaciones
- Bases biofísicas audiciónDocumento6 páginasBases biofísicas audiciónMaría MartínezAún no hay calificaciones
- Acústica 2023 T2Documento15 páginasAcústica 2023 T2luciaAún no hay calificaciones
- Manejo leche fresca plantas lácteasDocumento14 páginasManejo leche fresca plantas lácteasRutth Apolinario RicoAún no hay calificaciones
- Artículo GametogénesisDocumento9 páginasArtículo GametogénesisCecilia MendezAún no hay calificaciones
- PLAGASDocumento10 páginasPLAGASSHARONAún no hay calificaciones
- SENSOPERCEPCIÓNDocumento26 páginasSENSOPERCEPCIÓNJersiNaviaAún no hay calificaciones
- Ergonomía para La Industria MineraDocumento21 páginasErgonomía para La Industria MineraTrabajoenSaludAún no hay calificaciones
- Proyecto FinalDocumento19 páginasProyecto FinalyadiraAún no hay calificaciones
- Metodología Estándar de La Oms para El Análisis Del SemenDocumento44 páginasMetodología Estándar de La Oms para El Análisis Del Semenleidy100% (2)
- Libro Glaucoma Tomo 5Documento86 páginasLibro Glaucoma Tomo 5Hilda RenteriaAún no hay calificaciones
- Manejo Fitosanitario Mango PDFDocumento36 páginasManejo Fitosanitario Mango PDFAldo Beh MisAún no hay calificaciones
- Taller de Digestion. 123Documento18 páginasTaller de Digestion. 123dany luzAún no hay calificaciones
- Apraxias y Agnosias. RNNN Vol.15 (1), 2015Documento205 páginasApraxias y Agnosias. RNNN Vol.15 (1), 2015Camila Guerrero AravenaAún no hay calificaciones
- Ecto y Endoparasitos en BufalosDocumento11 páginasEcto y Endoparasitos en BufalosthaisAún no hay calificaciones
- TDAH y Su Relacion Con La Motivacion en El ContextDocumento7 páginasTDAH y Su Relacion Con La Motivacion en El ContextSel RivasAún no hay calificaciones
- Hoja de Seguridad 183-75Documento18 páginasHoja de Seguridad 183-75garcia.heberAún no hay calificaciones
- Alternativas Tecnologicas Que Permitan La Elaboracion de Productos Conformados Ricos en AG PoliinsatDocumento217 páginasAlternativas Tecnologicas Que Permitan La Elaboracion de Productos Conformados Ricos en AG PoliinsatMarion MarchettiAún no hay calificaciones
- PumaDocumento4 páginasPumaKike InoztrozaAún no hay calificaciones
- Practica No2 MicrosDocumento5 páginasPractica No2 MicrosANDRES BENAVIDESAún no hay calificaciones
- Carolina Stasiukynas Et Al. - 2021 - Las Carreteras Al Mar Estudio Sobre El Impacto deDocumento27 páginasCarolina Stasiukynas Et Al. - 2021 - Las Carreteras Al Mar Estudio Sobre El Impacto deJesus DurandAún no hay calificaciones
- El PrincipitoDocumento21 páginasEl PrincipitoprofesoresAún no hay calificaciones
- Botanica - Control de Malezas de Campo SucioDocumento3 páginasBotanica - Control de Malezas de Campo SucioDrodaAún no hay calificaciones
- EL ABORTO, Estudio BiblicoDocumento8 páginasEL ABORTO, Estudio BiblicoLuisMoraAún no hay calificaciones
- Bibliografia de Educacion AmbientalDocumento10 páginasBibliografia de Educacion Ambientaljose0% (1)
- Introducción Al Sistema Respiratorio y VentilaciónDocumento14 páginasIntroducción Al Sistema Respiratorio y VentilaciónNelson RangelAún no hay calificaciones
- Características y Cuidados Especiales para Pieles MadurasDocumento32 páginasCaracterísticas y Cuidados Especiales para Pieles MadurasSamantha DaniellaAún no hay calificaciones
- Medusas MísticasDocumento24 páginasMedusas MísticasAna Carolina DelgadoAún no hay calificaciones
- Sistema reproductor femenino y masculinoDocumento5 páginasSistema reproductor femenino y masculinoManuel HilarioAún no hay calificaciones
- El Croquis 162 - RCR ArquitectesDocumento143 páginasEl Croquis 162 - RCR ArquitectesAndrea Diaz Lacalle85% (13)
- Cebada PPDocumento44 páginasCebada PPAmos JoelAún no hay calificaciones
- Planeación de Clase Grado SegundoDocumento6 páginasPlaneación de Clase Grado SegundoGLORIA CECILIA GARCIA GARCIAAún no hay calificaciones
- Carboxy Min L MX WebDocumento1 páginaCarboxy Min L MX WebAndres AndradeAún no hay calificaciones
- La conciencia en el cerebro: Descifrando el enigma de cómo el cerebro elabora nuestros pensamientosDe EverandLa conciencia en el cerebro: Descifrando el enigma de cómo el cerebro elabora nuestros pensamientosCalificación: 2.5 de 5 estrellas2.5/5 (3)
- Cómo conversar con cualquiera: Descubre cómo ser la persona más interesante del lugar y a destruir la ansiedad social permanentementeDe EverandCómo conversar con cualquiera: Descubre cómo ser la persona más interesante del lugar y a destruir la ansiedad social permanentementeCalificación: 4 de 5 estrellas4/5 (81)
- Terapia cognitiva: Conceptos básicos y profundizaciónDe EverandTerapia cognitiva: Conceptos básicos y profundizaciónCalificación: 5 de 5 estrellas5/5 (1)
- Ansiedad infantil. Los trastornos explicados a los padresDe EverandAnsiedad infantil. Los trastornos explicados a los padresCalificación: 4.5 de 5 estrellas4.5/5 (25)
- Trauma, miedo y amor: Hacia una autonomía interior con la ayuda de las constelacionesDe EverandTrauma, miedo y amor: Hacia una autonomía interior con la ayuda de las constelacionesCalificación: 1 de 5 estrellas1/5 (1)
- Guía para superar los pensamientos atemorizantes, obsesivos o inquietantes: Libérate de los pensamientos negativos con la terapia cognitivo conductual (CBT)De EverandGuía para superar los pensamientos atemorizantes, obsesivos o inquietantes: Libérate de los pensamientos negativos con la terapia cognitivo conductual (CBT)Calificación: 4 de 5 estrellas4/5 (25)
- Resumen de Pensar rápido pensar despacio de Daniel KahnemanDe EverandResumen de Pensar rápido pensar despacio de Daniel KahnemanCalificación: 4.5 de 5 estrellas4.5/5 (11)
- Musicoterapia: Iniciación, técnicas y ejerciciosDe EverandMusicoterapia: Iniciación, técnicas y ejerciciosCalificación: 4.5 de 5 estrellas4.5/5 (13)
- Mente Organizada Cómo Pensar con Claridad y Hacer Todas Las Decisiones de Vida CorrectasDe EverandMente Organizada Cómo Pensar con Claridad y Hacer Todas Las Decisiones de Vida CorrectasCalificación: 4 de 5 estrellas4/5 (11)
- En busca de la mente: El largo camino para comprender la vida mental (y lo que aún queda por descubrir)De EverandEn busca de la mente: El largo camino para comprender la vida mental (y lo que aún queda por descubrir)Calificación: 4.5 de 5 estrellas4.5/5 (3)
- PNL: Programación neurolingüística: Una guía práctica y sencilla para iniciarse en la programación neurolingüísticaDe EverandPNL: Programación neurolingüística: Una guía práctica y sencilla para iniciarse en la programación neurolingüísticaCalificación: 4 de 5 estrellas4/5 (65)
- 50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaDe Everand50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaCalificación: 4 de 5 estrellas4/5 (37)
- Abrázame fuerte: Siete conversaciones para un amor duraderoDe EverandAbrázame fuerte: Siete conversaciones para un amor duraderoManu BerásteguiCalificación: 4.5 de 5 estrellas4.5/5 (13)
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)De EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Calificación: 5 de 5 estrellas5/5 (9)
- El autismo: Reflexiones y pautas para comprenderlo y abordarloDe EverandEl autismo: Reflexiones y pautas para comprenderlo y abordarloCalificación: 4 de 5 estrellas4/5 (7)
- DMT: La molécula del espíritu: Las revolucionarias investigaciones de un médico sobre la biología de las experiencias místicas y cercanas a la muerteDe EverandDMT: La molécula del espíritu: Las revolucionarias investigaciones de un médico sobre la biología de las experiencias místicas y cercanas a la muerteCalificación: 4 de 5 estrellas4/5 (135)
- El cerebro del niño explicado a los padresDe EverandEl cerebro del niño explicado a los padresCalificación: 4.5 de 5 estrellas4.5/5 (147)
- Cómo estimular el cerebro del niño: 100 ejercicios para potenciar la concentración, la memoria y otras funciones ejecutivasDe EverandCómo estimular el cerebro del niño: 100 ejercicios para potenciar la concentración, la memoria y otras funciones ejecutivasCalificación: 4.5 de 5 estrellas4.5/5 (12)
- Ansiedad: Vencer La Ansiedad, El Miedo, El Estrés Y La Depresión, Y Deshacerse De Las FobiasDe EverandAnsiedad: Vencer La Ansiedad, El Miedo, El Estrés Y La Depresión, Y Deshacerse De Las FobiasCalificación: 4.5 de 5 estrellas4.5/5 (17)
- Terapia cognitiva del comportamiento: Descubre cómo despejar tu cerebro con la TCC. Aprender a superar el miedo y la ansiedad, la depresiónDe EverandTerapia cognitiva del comportamiento: Descubre cómo despejar tu cerebro con la TCC. Aprender a superar el miedo y la ansiedad, la depresiónCalificación: 5 de 5 estrellas5/5 (3)
- Terapia de vidas pasadas: Un camino hacia la luz del alma. Técnica y prácticaDe EverandTerapia de vidas pasadas: Un camino hacia la luz del alma. Técnica y prácticaCalificación: 4.5 de 5 estrellas4.5/5 (11)
- Sesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesDe EverandSesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesCalificación: 4.5 de 5 estrellas4.5/5 (13)
- Psicología del Aprendizaje Humano: Adquisición de conocimiento y cambio personalDe EverandPsicología del Aprendizaje Humano: Adquisición de conocimiento y cambio personalCalificación: 4 de 5 estrellas4/5 (14)
- Psicoterapia breve con niños y adolescentesDe EverandPsicoterapia breve con niños y adolescentesCalificación: 4.5 de 5 estrellas4.5/5 (15)