También podría gustarte
- Ciclo de Krebs - BiologiaDocumento6 páginasCiclo de Krebs - BiologiaCARLA JOMBIAún no hay calificaciones
- Ciclo de KrebsDocumento12 páginasCiclo de KrebsGallardo RichardAún no hay calificaciones
- CICLO DE KREBS Power PointDocumento19 páginasCICLO DE KREBS Power PointAnibal Munguia67% (3)
- Examen - 6Documento3 páginasExamen - 6roger arieñ0% (2)
- Ciclo de Krebs MonografiaDocumento6 páginasCiclo de Krebs MonografiaRenato Chuquimbalqui Rivas100% (1)
- Ciclo de KrebsDocumento15 páginasCiclo de KrebsMiguel Angel Rodas Herrera100% (2)
- Ciclo de KrebsDocumento33 páginasCiclo de KrebsLN Brenda Janeth Beltrán DíazAún no hay calificaciones
- Etapas Ciclo KrebsDocumento10 páginasEtapas Ciclo KrebsCAMILABORAAún no hay calificaciones
- Guía MusescoreDocumento259 páginasGuía MusescoreDiego Sebastián García100% (1)
- DIA 1 SESION de APRENDIZAJE Elaboro Mi Propuesta de ActividadesDocumento6 páginasDIA 1 SESION de APRENDIZAJE Elaboro Mi Propuesta de Actividadesalexander alzamoraAún no hay calificaciones
- Resumen de Rutas Metabólicas BCMDocumento52 páginasResumen de Rutas Metabólicas BCMLuana HuertasAún no hay calificaciones
- Ciclo de KrebsDocumento19 páginasCiclo de KrebsWikii Wikoso0% (1)
- Ciclo de KrebsDocumento8 páginasCiclo de KrebsGustavo Adolfo MozAún no hay calificaciones
- Revolución IndustrialDocumento4 páginasRevolución IndustrialAlexander Núñez Cabarcas100% (3)
- 4to Boletin Semana 25-29 de JunioDocumento16 páginas4to Boletin Semana 25-29 de JunioWilber Ramos CartAún no hay calificaciones
- Introducción: Ciclo de KrebsDocumento5 páginasIntroducción: Ciclo de KrebsJuan David Losada RodriguezAún no hay calificaciones
- Ciclo de KrebsDocumento15 páginasCiclo de KrebsTania Lisset Oblea MejiaAún no hay calificaciones
- Ciclo de KrebsDocumento4 páginasCiclo de KrebsEirinuSanAún no hay calificaciones
- Ciclo de Krebs: Reacciones y producción de energía enDocumento5 páginasCiclo de Krebs: Reacciones y producción de energía enRanthal Rivadeneyra CastroAún no hay calificaciones
- CUESTIONARIODocumento8 páginasCUESTIONARIOJuan Jose Baros AyalaAún no hay calificaciones
- Colegio Adventista PimentelDocumento26 páginasColegio Adventista PimentelCamila RubioAún no hay calificaciones
- APUNTESDocumento4 páginasAPUNTESFroylan MezaAún no hay calificaciones
- Ciclo de Krebs: ruta metabólica claveDocumento6 páginasCiclo de Krebs: ruta metabólica claveDiianaLauraMelendezAún no hay calificaciones
- Ciclo de Krebs BioquimicaDocumento8 páginasCiclo de Krebs BioquimicaYeremi MinauroAún no hay calificaciones
- El Ciclo de KrebsDocumento8 páginasEl Ciclo de KrebsTlacaélel Rafael Cáceres Santa CruzAún no hay calificaciones
- Ciclo de KrebsDocumento13 páginasCiclo de KrebsCRISTHIAN GONZALES CATAYAún no hay calificaciones
- Ciclo KrebsDocumento13 páginasCiclo KrebsAlex Jimeno0% (1)
- Ciclo de KrebsDocumento14 páginasCiclo de KrebsOscar TrujilloAún no hay calificaciones
- Exposicion Ciclo de KrebsDocumento6 páginasExposicion Ciclo de KrebsJesús Rafael Salazar MendozaAún no hay calificaciones
- Ciclo de KlebsDocumento9 páginasCiclo de KlebsIram Fernando Oleta CruzAún no hay calificaciones
- Laura BioquimicaDocumento19 páginasLaura BioquimicaLauren AmayaAún no hay calificaciones
- Ciclo de KrebsDocumento33 páginasCiclo de KrebsJosé Álvarez RoblesAún no hay calificaciones
- Artículo Del Ciclo de Krebs Mayo 31 2018Documento5 páginasArtículo Del Ciclo de Krebs Mayo 31 2018Lesdy MarllyAún no hay calificaciones
- Ciclo Krebs resumenDocumento6 páginasCiclo Krebs resumenJose Luis Arroyo Hernandez0% (1)
- El Ciclo de KrebsDocumento2 páginasEl Ciclo de Krebs0532010015 JOAS SAMUEL VEGA SERPA ESTUDIANTE ACTIVOAún no hay calificaciones
- Ciclo de Krebs 22Documento11 páginasCiclo de Krebs 22Mari CastedoAún no hay calificaciones
- Ciclo de Krebs y El MetabolismoDocumento7 páginasCiclo de Krebs y El Metabolismoirvin reyes rosalesAún no hay calificaciones
- Ciclo de Krebs BasicoDocumento9 páginasCiclo de Krebs BasicoJavier AranaAún no hay calificaciones
- El ciclo de Krebs: reacciones y productosDocumento3 páginasEl ciclo de Krebs: reacciones y productosɆsƬeŁa尺Aún no hay calificaciones
- Metabolitos IntermediariosDocumento3 páginasMetabolitos IntermediariosFelipe ChimborazoAún no hay calificaciones
- Ciclo de Krebs. Grupo 4. Bioquímica II.Documento11 páginasCiclo de Krebs. Grupo 4. Bioquímica II.Zuly AGomezAún no hay calificaciones
- Metabolismo de carbohidratos: glucólisis, ciclo de Krebs y cadena respiratoriaDocumento15 páginasMetabolismo de carbohidratos: glucólisis, ciclo de Krebs y cadena respiratoriaCarolay ParedesAún no hay calificaciones
- Resumen 12 Ciclo de KrebsDocumento12 páginasResumen 12 Ciclo de KrebsMariam Cabellos SAún no hay calificaciones
- Ciclo de Krebs: reacciones y producción de ATPDocumento11 páginasCiclo de Krebs: reacciones y producción de ATPjuan pabloAún no hay calificaciones
- Ciclo de KerbsDocumento4 páginasCiclo de KerbsÁngel Archila100% (1)
- 2.1. TAREA Prác. Ciclo de KrebsDocumento8 páginas2.1. TAREA Prác. Ciclo de KrebsBAZÁN SALDAÑA DULCE VICTORIAAún no hay calificaciones
- Ciclo de Krebs TutoríaDocumento2 páginasCiclo de Krebs TutoríaBeverlyAún no hay calificaciones
- El Ciclo de KrebsDocumento20 páginasEl Ciclo de KrebsKotetu369Aún no hay calificaciones
- Ciclo de Krebs Seminario 2dauDocumento8 páginasCiclo de Krebs Seminario 2dauDanixaKristelBarriosUcañayAún no hay calificaciones
- Piruvato DeshidrogenasaDocumento7 páginasPiruvato DeshidrogenasaRoberth Daniel Torres GutierrezAún no hay calificaciones
- Ciclo de KrebsDocumento13 páginasCiclo de KrebsDiaz BrunoAún no hay calificaciones
- Ciclo Del Acido CitricoDocumento12 páginasCiclo Del Acido CitricoROXANAAún no hay calificaciones
- Ciclo de KrebsDocumento5 páginasCiclo de KrebsCesarAún no hay calificaciones
- Ciclo Del Ácido Cítrico y GlucolisisDocumento18 páginasCiclo Del Ácido Cítrico y GlucolisisJerry Carhuatanta SaavedraAún no hay calificaciones
- Ciclo de Krebs: Resumen del proceso metabólicoDocumento7 páginasCiclo de Krebs: Resumen del proceso metabólicoRenzo GonzalezAún no hay calificaciones
- CicloKrebs38ATPDocumento5 páginasCicloKrebs38ATPMaruUzcateguiReinozaAún no hay calificaciones
- El Ciclo de KrebsDocumento1 páginaEl Ciclo de KrebsZulema GaitánAún no hay calificaciones
- CicloK GeorgetteDocumento6 páginasCicloK Georgetteosmigdy mataAún no hay calificaciones
- Ciclo de KrebsDocumento10 páginasCiclo de KrebsEliannys Arregoces BritoAún no hay calificaciones
- Ciclo de Krebs - Exposición de F. de Micro y BioDocumento29 páginasCiclo de Krebs - Exposición de F. de Micro y BioMaria Jose QuirozAún no hay calificaciones
- Ciclo de Krebs: Visión general del metabolismo celularDocumento14 páginasCiclo de Krebs: Visión general del metabolismo celularmay_street_von100% (1)
- Ciclo de Krebs o Ciclo Del Ácido CítricoDocumento5 páginasCiclo de Krebs o Ciclo Del Ácido CítricoGaby RodriguezAún no hay calificaciones
- Ciclo de Krebs Bioquimica Carla BislipDocumento4 páginasCiclo de Krebs Bioquimica Carla BislipcarlaAún no hay calificaciones
- Ciclo de KrebsDocumento16 páginasCiclo de KrebsCarmen Flores100% (4)
- Ponchar rj45Documento19 páginasPonchar rj45RamonValdesAún no hay calificaciones
- Grupo de Trabajo de Escenografia OfficeDocumento1 páginaGrupo de Trabajo de Escenografia OfficeEl anonimo oscuroAún no hay calificaciones
- Eda 3Documento8 páginasEda 3El anonimo oscuroAún no hay calificaciones
- Ferti 2023 - 025659Documento14 páginasFerti 2023 - 025659El anonimo oscuroAún no hay calificaciones
- YhosiDocumento39 páginasYhosiEl anonimo oscuroAún no hay calificaciones
- Angel Corregido 2.Documento1 páginaAngel Corregido 2.El anonimo oscuroAún no hay calificaciones
- Modulo LombriDocumento20 páginasModulo LombriEl anonimo oscuroAún no hay calificaciones
- Informe Experimental CristianDocumento2 páginasInforme Experimental CristianEl anonimo oscuroAún no hay calificaciones
- Informe Experimental Cristian.Documento2 páginasInforme Experimental Cristian.El anonimo oscuroAún no hay calificaciones
- Guía para Determinar La Dap y PorosidadDocumento5 páginasGuía para Determinar La Dap y PorosidadEl anonimo oscuroAún no hay calificaciones
- Ruben Fertilodad 125408Documento15 páginasRuben Fertilodad 125408El anonimo oscuroAún no hay calificaciones
- Valles y AltiplanoDocumento26 páginasValles y AltiplanoEl anonimo oscuroAún no hay calificaciones
- Informe de Economía Agrícola (Factor Capital) - 1Documento9 páginasInforme de Economía Agrícola (Factor Capital) - 1El anonimo oscuroAún no hay calificaciones
- 23 Entomologia Agrícola - PLANILLAS DE EVALUACIÓN CONTINUADocumento2 páginas23 Entomologia Agrícola - PLANILLAS DE EVALUACIÓN CONTINUAEl anonimo oscuroAún no hay calificaciones
- Informe de Aforo de Caudal111 - 061739Documento4 páginasInforme de Aforo de Caudal111 - 061739El anonimo oscuroAún no hay calificaciones
- Los Dominios de Las Paginas Web PDFDocumento3 páginasLos Dominios de Las Paginas Web PDFmary garcesAún no hay calificaciones
- GUIA 11 DETERMINACION DE PARAMETROS EDAFICOS ESTUDIADOS EN UNA CALICATA - RemovedDocumento15 páginasGUIA 11 DETERMINACION DE PARAMETROS EDAFICOS ESTUDIADOS EN UNA CALICATA - RemovedjngavilanesAún no hay calificaciones
- Preguntas H1Documento92 páginasPreguntas H1Alejandro MarínAún no hay calificaciones
- Copia de CONTROL DE LECTURADocumento2 páginasCopia de CONTROL DE LECTURAJavier Alejandro Lara CorreaAún no hay calificaciones
- PRACTICA EDUCATIVA II Programas UNEFA Licenciatura Educación Integral Octavo Semestre 8vo. SemestreDocumento5 páginasPRACTICA EDUCATIVA II Programas UNEFA Licenciatura Educación Integral Octavo Semestre 8vo. SemestreJhony PradoAún no hay calificaciones
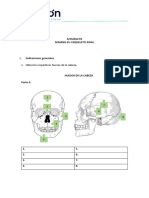
- Tarea Esqueleto AxialDocumento6 páginasTarea Esqueleto AxialdieneAún no hay calificaciones
- AHUMADODocumento42 páginasAHUMADOXimena Villanueva CastilloAún no hay calificaciones
- Equipos ATEXDocumento31 páginasEquipos ATEXRoberto ZabalaAún no hay calificaciones
- Mediciones de laboratorioDocumento18 páginasMediciones de laboratoriojosezxcAún no hay calificaciones
- Conta de Gerencia Preguntas LaboratorioDocumento3 páginasConta de Gerencia Preguntas LaboratorioPaul Dalhy ZapataAún no hay calificaciones
- Grado 6 ArtísticaDocumento2 páginasGrado 6 ArtísticaUnidad Educación UGAún no hay calificaciones
- Deóntica JuridicaDocumento10 páginasDeóntica JuridicapegabesaAún no hay calificaciones
- ObjetivoDocumento5 páginasObjetivoKarla Dayana AriasAún no hay calificaciones
- Universidad Abierta Y A Distancia de Mexico Docente: Lic. Liliana Lopez AstorgaDocumento4 páginasUniversidad Abierta Y A Distancia de Mexico Docente: Lic. Liliana Lopez AstorgaMhoni GarciaAún no hay calificaciones
- Tema 7 H CuantitativaDocumento37 páginasTema 7 H CuantitativaDan Chate Velasque.Aún no hay calificaciones
- Riesgos financieros en la Universidad Austral del CuscoDocumento21 páginasRiesgos financieros en la Universidad Austral del CuscoElio Lazo Chayan0% (1)
- Elación de Pertenencia Y No Pertenencia: ActividadesDocumento6 páginasElación de Pertenencia Y No Pertenencia: ActividadesAfulitoAún no hay calificaciones
- Apunte 11. Dimensionamiento de Pavimentos FlexiblesDocumento264 páginasApunte 11. Dimensionamiento de Pavimentos FlexiblesJhonny Gabriel TorrezAún no hay calificaciones
- TEOLOGIADocumento3 páginasTEOLOGIAHernandez LokilloAún no hay calificaciones
- WD 3227SP 04.inddDocumento1 páginaWD 3227SP 04.inddCeleynes RTAún no hay calificaciones
- Procesos Metalurgicos Laboratorio N 2 inDocumento18 páginasProcesos Metalurgicos Laboratorio N 2 inHeydi CalloAún no hay calificaciones
- Guía de 7 pasos para aprender melódica enDocumento15 páginasGuía de 7 pasos para aprender melódica enSustentando LA Vida100% (1)
- Fonetica Conceptos (Escrito Por Mi)Documento10 páginasFonetica Conceptos (Escrito Por Mi)alexroglaAún no hay calificaciones
- El Hongo Kombucha: Summary. ILLANA, C. (2007) - The Fungi Kombucha. Bol. Soc. Micol. Madrid 31: 269-272Documento4 páginasEl Hongo Kombucha: Summary. ILLANA, C. (2007) - The Fungi Kombucha. Bol. Soc. Micol. Madrid 31: 269-272leonel perezAún no hay calificaciones
- Determinacion Del Requerimiento Hidrico Del Cultivo de LechugaDocumento6 páginasDeterminacion Del Requerimiento Hidrico Del Cultivo de LechugaMICHELLE STEFANY CAIZA CLAVIJOAún no hay calificaciones