También podría gustarte
- Alteración y Tipos de Alteración en AlimentosDocumento8 páginasAlteración y Tipos de Alteración en AlimentosEliseo QuispeAún no hay calificaciones
- Preservacion Dealimentos - QuinuaDocumento29 páginasPreservacion Dealimentos - QuinuaEliseo QuispeAún no hay calificaciones
- Perspectivas futuras para las tecnologías de enzimas en la industria alimentariaDocumento10 páginasPerspectivas futuras para las tecnologías de enzimas en la industria alimentariaEliseo QuispeAún no hay calificaciones
- Grupo de Problema 1Documento2 páginasGrupo de Problema 1Eliseo QuispeAún no hay calificaciones
- Presentació N Investigacion Cientifica Yogurt de Diente de LeonDocumento32 páginasPresentació N Investigacion Cientifica Yogurt de Diente de LeonEliseo QuispeAún no hay calificaciones
- Examen Trig TrilceDocumento1 páginaExamen Trig TrilceEliseo QuispeAún no hay calificaciones
- Examen AlgebraDocumento1 páginaExamen AlgebraEliseo QuispeAún no hay calificaciones
- Trigonometría básica privada TrilceDocumento1 páginaTrigonometría básica privada TrilceEliseo QuispeAún no hay calificaciones
- Yogur diente león ayuda hígado pielDocumento1 páginaYogur diente león ayuda hígado pielileanaAún no hay calificaciones
- Medimos el perímetro de polígonosDocumento2 páginasMedimos el perímetro de polígonosEliseo QuispeAún no hay calificaciones
- Examen ARITMETICADocumento2 páginasExamen ARITMETICAEliseo QuispeAún no hay calificaciones
- Examen aRITMETICA 2Documento1 páginaExamen aRITMETICA 2Eliseo QuispeAún no hay calificaciones
- Triptico TunsumaDocumento2 páginasTriptico TunsumaEliseo QuispeAún no hay calificaciones
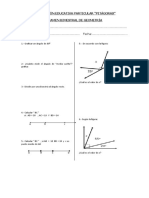
- Examen bimestral de geometría del colegio PitágorasDocumento1 páginaExamen bimestral de geometría del colegio PitágorasEliseo QuispeAún no hay calificaciones
- Cartel de Competencias y Capacidades Desempeños PrimariaDocumento33 páginasCartel de Competencias y Capacidades Desempeños PrimariaEliseo QuispeAún no hay calificaciones
- SESIÓN SustantivosDocumento3 páginasSESIÓN SustantivosEliseo QuispeAún no hay calificaciones
- DIPLOMA Kisio 2018Documento1 páginaDIPLOMA Kisio 2018Eliseo QuispeAún no hay calificaciones
- Ficha COMUNICACIÓN 5° OKDocumento9 páginasFicha COMUNICACIÓN 5° OKEliseo QuispeAún no hay calificaciones
- Cuadro Nominal de Asignación Personal Docente de La IDocumento2 páginasCuadro Nominal de Asignación Personal Docente de La IEliseo QuispeAún no hay calificaciones
- Ficha Cyt ManiferosDocumento1 páginaFicha Cyt ManiferosEliseo QuispeAún no hay calificaciones
- Ficha de Inscripción JDEN 2022Documento6 páginasFicha de Inscripción JDEN 2022Eliseo QuispeAún no hay calificaciones
- Ficha C y T El AguaDocumento4 páginasFicha C y T El AguaEliseo QuispeAún no hay calificaciones
- TRIPTICO2021 SDocumento3 páginasTRIPTICO2021 SEliseo QuispeAún no hay calificaciones
- Ficha El ArticuloDocumento3 páginasFicha El ArticuloEliseo QuispeAún no hay calificaciones
- Ficha C y T El AireDocumento3 páginasFicha C y T El AireEliseo QuispeAún no hay calificaciones
- Ficha Comunicacion Setiembre 2Documento4 páginasFicha Comunicacion Setiembre 2Eliseo QuispeAún no hay calificaciones
- Ficha Clases de ConjuntosDocumento4 páginasFicha Clases de ConjuntosEliseo QuispeAún no hay calificaciones
- Ficha Personal Social AgostoDocumento3 páginasFicha Personal Social AgostoEliseo QuispeAún no hay calificaciones
- Unidad2 Sesion1Documento24 páginasUnidad2 Sesion1Zeus Cronos VillarrealAún no hay calificaciones
- Listin de Cds Ingenieria ElectronicaDocumento920 páginasListin de Cds Ingenieria ElectronicarobertoAún no hay calificaciones
- Cuestionario 5 PDFDocumento3 páginasCuestionario 5 PDFRonald Capo0% (1)
- Estequiometria Calculo Masa A MasaDocumento1 páginaEstequiometria Calculo Masa A MasaRudy100% (1)
- 4 PDFDocumento10 páginas4 PDFJavier Saul Gonzalez ReyesAún no hay calificaciones
- Humidificación 2Documento69 páginasHumidificación 2Majo27Aún no hay calificaciones
- NC 220-4-2006 Eficienc Energ EdificacDocumento11 páginasNC 220-4-2006 Eficienc Energ EdificacDescargar scribdAún no hay calificaciones
- Guia Autoaprendizaje Estudiante 2do Bto Ciencia f1 s3Documento6 páginasGuia Autoaprendizaje Estudiante 2do Bto Ciencia f1 s3Mauro Bermudez CaballieriAún no hay calificaciones
- Asnos EstupidosDocumento2 páginasAsnos EstupidosDanielaBurgos0% (2)
- Tecnología del zincDocumento115 páginasTecnología del zincJuve NiflaAún no hay calificaciones
- Planificación Modelo DUA Por Unidad 2019 8 BásicoDocumento5 páginasPlanificación Modelo DUA Por Unidad 2019 8 BásicoJulio Matías Salas NorambuenaAún no hay calificaciones
- Curso de PR para Dirigir Instalaciones de Rayos XDocumento261 páginasCurso de PR para Dirigir Instalaciones de Rayos XLazaro A. Moynelo DiazAún no hay calificaciones
- Afectaciones Del ConcretoDocumento3 páginasAfectaciones Del ConcretoLuisaFuentesAún no hay calificaciones
- AVANCE DE TESIS 50% Corregido 4Documento41 páginasAVANCE DE TESIS 50% Corregido 4Luis HuAún no hay calificaciones
- Tycho BraheDocumento11 páginasTycho BraheJulio CesarAún no hay calificaciones
- Contacto Directo de Dos Fases InmisciblesDocumento4 páginasContacto Directo de Dos Fases InmisciblesVanesa Chugñay33% (3)
- Taller ClimatologiaDocumento22 páginasTaller ClimatologiaErika Cruz50% (2)
- Evau RedoxDocumento14 páginasEvau RedoxIsabelMontillaArteagaAún no hay calificaciones
- Sesion 6: Principios de Los de Los Procesos Cíclicos Mquinas Térmicas y Su RendimientoDocumento28 páginasSesion 6: Principios de Los de Los Procesos Cíclicos Mquinas Térmicas y Su RendimientoANA LELIS BELINDA QUENTA QUINOAún no hay calificaciones
- La Tabla PeriodicaDocumento44 páginasLa Tabla Periodicamaria silviaAún no hay calificaciones
- T3P2 - Carlos Alexis Prieto Llamas 19300352 7ADocumento24 páginasT3P2 - Carlos Alexis Prieto Llamas 19300352 7ACARLOS ALEXIS PRIETO LLAMASAún no hay calificaciones
- FT Aw Ferronilox 60Documento2 páginasFT Aw Ferronilox 60franklinAún no hay calificaciones
- Informe 2 Terminarrrrr Ope 2Documento24 páginasInforme 2 Terminarrrrr Ope 2Cruz Rojas CristinaAún no hay calificaciones
- Matpel Nivel I (Manual Completo)Documento107 páginasMatpel Nivel I (Manual Completo)Julio OrjedaAún no hay calificaciones
- Propiedades comparativas de candelas de cera de abeja y parafinaDocumento19 páginasPropiedades comparativas de candelas de cera de abeja y parafinaJessica HerreraAún no hay calificaciones
- 2 Far Maco Cine TicaDocumento190 páginas2 Far Maco Cine TicaJuan FedericoAún no hay calificaciones
- Capitulo - 1 Materia y MedicionDocumento64 páginasCapitulo - 1 Materia y MedicionDIEGO A PAGAN-BARRETOAún no hay calificaciones
- Inducción Correct Ucv11062020185513Documento23 páginasInducción Correct Ucv11062020185513VICTOR CAMPOS DONOSOAún no hay calificaciones
- Trabajo reversible vs irreversibleDocumento2 páginasTrabajo reversible vs irreversiblerafaelAún no hay calificaciones
- Fundamentos de La Fundicion de Metales TDocumento9 páginasFundamentos de La Fundicion de Metales TBenito Merida FloresAún no hay calificaciones
- Informe 1 Reactor de Nucleo de HierroDocumento32 páginasInforme 1 Reactor de Nucleo de HierroHerberth Villachica CarranzaAún no hay calificaciones
- Influencia. La psicología de la persuasiónDe EverandInfluencia. La psicología de la persuasiónCalificación: 4.5 de 5 estrellas4.5/5 (14)
- Desastres y emergencias. Prevención, mitigación y preparaciónDe EverandDesastres y emergencias. Prevención, mitigación y preparaciónCalificación: 4 de 5 estrellas4/5 (5)
- La revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaDe EverandLa revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaCalificación: 5 de 5 estrellas5/5 (200)
- Disciplina Mental: Técnicas infalibles para lograr todo lo que te propones y eliminar la pereza y la procrastinación de tu vida para siempreDe EverandDisciplina Mental: Técnicas infalibles para lograr todo lo que te propones y eliminar la pereza y la procrastinación de tu vida para siempreCalificación: 5 de 5 estrellas5/5 (3)
- Neurociencia para vencer la depresión: La esprial ascendenteDe EverandNeurociencia para vencer la depresión: La esprial ascendenteCalificación: 4.5 de 5 estrellas4.5/5 (10)
- Clics contra la humanidad: Libertad y resistencia en la era de la distracción tecnológicaDe EverandClics contra la humanidad: Libertad y resistencia en la era de la distracción tecnológicaCalificación: 4.5 de 5 estrellas4.5/5 (117)
- Anatomía & 100 estiramientos Esenciales (Color): Técnicas, beneficios, precauciones, consejos, tablas de series, dolenciasDe EverandAnatomía & 100 estiramientos Esenciales (Color): Técnicas, beneficios, precauciones, consejos, tablas de series, dolenciasCalificación: 4.5 de 5 estrellas4.5/5 (21)
- Interpretación del ECG: Una Guía Práctica e Intuitiva para Aprender a Leer el ECG y Diagnosticar y Tratar ArritmiasDe EverandInterpretación del ECG: Una Guía Práctica e Intuitiva para Aprender a Leer el ECG y Diagnosticar y Tratar ArritmiasAún no hay calificaciones
- La metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceDe EverandLa metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceCalificación: 5 de 5 estrellas5/5 (8)
- Trucos para Leer la Mente de los Demás: Cómo Adivinar el Pensamiento de los Demás con Poco Esfuerzo. 2 Libros en 1 - Secretos de la Psicología Oscura, Cómo ser un Detector de MentirasDe EverandTrucos para Leer la Mente de los Demás: Cómo Adivinar el Pensamiento de los Demás con Poco Esfuerzo. 2 Libros en 1 - Secretos de la Psicología Oscura, Cómo ser un Detector de MentirasCalificación: 4.5 de 5 estrellas4.5/5 (3)
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- Reconstrucción de dientes endodonciados: Pautas de actuación clínicaDe EverandReconstrucción de dientes endodonciados: Pautas de actuación clínicaCalificación: 5 de 5 estrellas5/5 (4)
- Zensorialmente : Dejá que tu cuerpo sea tu cerebroDe EverandZensorialmente : Dejá que tu cuerpo sea tu cerebroAún no hay calificaciones
- El libro conciso de la punción seca: Manual del terapeuta para las aplicaciones en los puntos gatillo miofasciales (Color)De EverandEl libro conciso de la punción seca: Manual del terapeuta para las aplicaciones en los puntos gatillo miofasciales (Color)Calificación: 3 de 5 estrellas3/5 (2)
- Fisiopatología de las enfermedades cardiovascularesDe EverandFisiopatología de las enfermedades cardiovascularesCalificación: 5 de 5 estrellas5/5 (1)
- Nuevo manual de gastronomía molecular: El encuentro entre la ciencia y la cocinaDe EverandNuevo manual de gastronomía molecular: El encuentro entre la ciencia y la cocinaCalificación: 4.5 de 5 estrellas4.5/5 (4)
- LAS VELAS JAPONESAS DE UNA FORMA SENCILLA. La guía de introducción a las velas japonesas y a las estrategias de análisis técnico más eficaces.De EverandLAS VELAS JAPONESAS DE UNA FORMA SENCILLA. La guía de introducción a las velas japonesas y a las estrategias de análisis técnico más eficaces.Calificación: 4.5 de 5 estrellas4.5/5 (54)
- 7 tendencias digitales que cambiarán el mundoDe Everand7 tendencias digitales que cambiarán el mundoCalificación: 4.5 de 5 estrellas4.5/5 (87)
- Excel para principiantes: Aprenda a utilizar Excel 2016, incluyendo una introducción a fórmulas, funciones, gráficos, cuadros, macros, modelado, informes, estadísticas, Excel Power Query y másDe EverandExcel para principiantes: Aprenda a utilizar Excel 2016, incluyendo una introducción a fórmulas, funciones, gráficos, cuadros, macros, modelado, informes, estadísticas, Excel Power Query y másCalificación: 2.5 de 5 estrellas2.5/5 (3)
- Notas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesDe EverandNotas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesAún no hay calificaciones
- Borges y la memoria: De "Funes el memorioso" a la neurona de Jennifer AnistonDe EverandBorges y la memoria: De "Funes el memorioso" a la neurona de Jennifer AnistonCalificación: 5 de 5 estrellas5/5 (4)
- Receta De La Freidora De Aire Libro De Cocina De La Freidora De Aire/ Air Fryer Cookbook Spanish VersionDe EverandReceta De La Freidora De Aire Libro De Cocina De La Freidora De Aire/ Air Fryer Cookbook Spanish VersionCalificación: 4.5 de 5 estrellas4.5/5 (2)