También podría gustarte
- 38 6nnbecquerDocumento17 páginas38 6nnbecquerbecquer tuctoAún no hay calificaciones
- Microalgas y La Economia CircularDocumento8 páginasMicroalgas y La Economia CircularJesús Eduardo De la CruzAún no hay calificaciones
- Técnicas de Muestreo y Preservado de Algas Marinas (Microalgas, Macroalgas y Plantas Vasculares Marinas)Documento20 páginasTécnicas de Muestreo y Preservado de Algas Marinas (Microalgas, Macroalgas y Plantas Vasculares Marinas)javi jimenezAún no hay calificaciones
- El Microfitoplancton en Los PrincipalesDocumento15 páginasEl Microfitoplancton en Los PrincipalesJuan David SantanaAún no hay calificaciones
- Riesgo Biologico Asociado Al Tratamiento de Aguas ResidualesDocumento5 páginasRiesgo Biologico Asociado Al Tratamiento de Aguas ResidualesCesar GrijalvaAún no hay calificaciones
- Purificacion de Agua Del Rio Chicama Por Biorremediacion Con Chondracanthus Chamissoi 1Documento45 páginasPurificacion de Agua Del Rio Chicama Por Biorremediacion Con Chondracanthus Chamissoi 1Ignacio CamachoAún no hay calificaciones
- Rev Colombiana - Caleta OliviaDocumento13 páginasRev Colombiana - Caleta OliviaGraciela PucciAún no hay calificaciones
- FITOPLANCTONDocumento11 páginasFITOPLANCTONCristian Basilik CasasAún no hay calificaciones
- Biorremediación Con Microalgas para Combatir La EutrofizaciónDocumento4 páginasBiorremediación Con Microalgas para Combatir La EutrofizaciónCamii SiilvaAún no hay calificaciones
- Sistema de Acuaponía, Peces. Tomo 6: Sistemas de acuaponíaDe EverandSistema de Acuaponía, Peces. Tomo 6: Sistemas de acuaponíaAún no hay calificaciones
- Textodelartculo 144814505351 1 10 20230603Documento21 páginasTextodelartculo 144814505351 1 10 20230603Beatriz López SánchezAún no hay calificaciones
- Diagnostico de La Calidad Del Agua en Sistemas Loticos Utilizando Diatomeas y MacroinvertebradosDocumento11 páginasDiagnostico de La Calidad Del Agua en Sistemas Loticos Utilizando Diatomeas y MacroinvertebradosGabo MontesAún no hay calificaciones
- Sistema de acuaponía, peces. Tomo 3: Sistemas de acuaponíaDe EverandSistema de acuaponía, peces. Tomo 3: Sistemas de acuaponíaAún no hay calificaciones
- Okeanos - BiomarcadoresDocumento5 páginasOkeanos - BiomarcadoresAlicia Herrera UlibarriAún no hay calificaciones
- Ejemplo de Un Trahjo de Investigaciontrabajo-D-MDocumento13 páginasEjemplo de Un Trahjo de Investigaciontrabajo-D-MAldo Ezequilla RamirezAún no hay calificaciones
- CianobacteriasDocumento5 páginasCianobacteriasMisael ArauzAún no hay calificaciones
- Metagenomic 2019Documento24 páginasMetagenomic 2019Arturo LiñanAún no hay calificaciones
- Lectura y Analisis de ArticulosDocumento3 páginasLectura y Analisis de ArticulosJose CumbicosAún no hay calificaciones
- El Uso de Helmintos Parásitos Como Bioindicadores en La Evaluación de La Calidad Del Agua MexicoDocumento12 páginasEl Uso de Helmintos Parásitos Como Bioindicadores en La Evaluación de La Calidad Del Agua MexicoElizaAún no hay calificaciones
- Hidrobiológica 0188-8897: Issn: Rehb@xanum - Uam.mxDocumento15 páginasHidrobiológica 0188-8897: Issn: Rehb@xanum - Uam.mxareamedioambientebegasAún no hay calificaciones
- Membranas Nano PDFDocumento15 páginasMembranas Nano PDFLindaNathalieSuarezPiñerezAún no hay calificaciones
- Macroinvertebrados Como Bioindicadores Del Río PiuraDocumento8 páginasMacroinvertebrados Como Bioindicadores Del Río PiuradcoronelmedranoAún no hay calificaciones
- C9-Microorganismos de Ecosistemas Acuáticos-MaDocumento20 páginasC9-Microorganismos de Ecosistemas Acuáticos-MaVEGA MACAS PIERO JUNIORAún no hay calificaciones
- Resumen Protocolo de Investigacion.Documento4 páginasResumen Protocolo de Investigacion.yaneliAún no hay calificaciones
- Efecto Del Fotoperiodo Sobre Chaetocero Calcitrans-Villa, Herazo y Torregroza, 2014Documento7 páginasEfecto Del Fotoperiodo Sobre Chaetocero Calcitrans-Villa, Herazo y Torregroza, 2014altamar20Aún no hay calificaciones
- Biotecnologia en Colores (1.1)Documento10 páginasBiotecnologia en Colores (1.1)stevenAún no hay calificaciones
- 11 ABV2021-11LopezycolDocumento11 páginas11 ABV2021-11LopezycolBeatriz López SánchezAún no hay calificaciones
- Potencial Biotecnológico de Los Hongos Marinos en Las Zonas Costeras de Costa RicaDocumento12 páginasPotencial Biotecnológico de Los Hongos Marinos en Las Zonas Costeras de Costa RicaLuis Ariel Beristain AguirreAún no hay calificaciones
- 4 InsecticidasselectivosDocumento12 páginas4 InsecticidasselectivosJesusPJAún no hay calificaciones
- 950 1498 1 SMDocumento7 páginas950 1498 1 SMIrene SilvaAún no hay calificaciones
- Coliformes Fecales Agua-Sedimentos PDFDocumento10 páginasColiformes Fecales Agua-Sedimentos PDFCarlos MuñozAún no hay calificaciones
- Leyte Manriqueetal2022Documento13 páginasLeyte Manriqueetal2022AlekkerAún no hay calificaciones
- SadebaclobosDocumento3 páginasSadebaclobosAlejo Garcia SeguiAún no hay calificaciones
- Articulo Publicado Calidad Bacteriológica de Las Piscinas de La Ciudad de Jaén, 2022 - Ciencia Latina Revista Científica MultidisciplinarDocumento13 páginasArticulo Publicado Calidad Bacteriológica de Las Piscinas de La Ciudad de Jaén, 2022 - Ciencia Latina Revista Científica MultidisciplinarALEXANDER ALDAIR PEREZ NAVARROAún no hay calificaciones
- Estudio de Farmacos ContaminatesDocumento29 páginasEstudio de Farmacos ContaminatesMiguel BERRIOSAún no hay calificaciones
- Toledo y Razo 2018Documento6 páginasToledo y Razo 2018frfunkAún no hay calificaciones
- Admin,+art +1Documento10 páginasAdmin,+art +1oerstedella1Aún no hay calificaciones
- Cultivo de BioflocDocumento9 páginasCultivo de BioflocsemiconductormanAún no hay calificaciones
- 5077-Texto Del Artículo-40487-1-10-20220524Documento10 páginas5077-Texto Del Artículo-40487-1-10-20220524ANGEL RAMON MACIAS RODRIGUEZAún no hay calificaciones
- Aplicaciones de La BacteriologiaDocumento4 páginasAplicaciones de La BacteriologiaEduardo Rodriguez0% (1)
- Informe Completo de Mortalidad de TortugasDocumento16 páginasInforme Completo de Mortalidad de TortugasConclusión Diario DigitalAún no hay calificaciones
- Informe-Sistema Producción AcuícolaDocumento28 páginasInforme-Sistema Producción AcuícolaLeslye Lorena Quiroga MoratoAún no hay calificaciones
- Articulo de Venezuela-ChiflesDocumento16 páginasArticulo de Venezuela-ChiflesValeria PovedaAún no hay calificaciones
- BiosensoresDocumento22 páginasBiosensoresVanessa Bermello JiménezAún no hay calificaciones
- Utilización de Microorganismos de Ambientes ExtremosDocumento13 páginasUtilización de Microorganismos de Ambientes ExtremosFertilizantes FosyeikiAún no hay calificaciones
- Mucus en CoralesDocumento8 páginasMucus en CoralesIvanAntonioVilchezAún no hay calificaciones
- MarinaDocumento181 páginasMarinaLeonel HerreraAún no hay calificaciones
- 2293-Texto Del Artículo-11458-1-10-20220511Documento8 páginas2293-Texto Del Artículo-11458-1-10-20220511Eric GalindoAún no hay calificaciones
- ABV Vol 42 (1) 2022 ContenidoDocumento7 páginasABV Vol 42 (1) 2022 ContenidoAmilcar FerminAún no hay calificaciones
- 1586-Texto Del Artículo-11102-2-10-20221206Documento6 páginas1586-Texto Del Artículo-11102-2-10-20221206StelleAún no hay calificaciones
- Tecnologías para El Cultivo de Microalgas en Interior y Exterior. Propuestas Prácticas para Operar en Granjas Acuícolas de Pequeña EscalaDocumento13 páginasTecnologías para El Cultivo de Microalgas en Interior y Exterior. Propuestas Prácticas para Operar en Granjas Acuícolas de Pequeña EscalaLuis Fernando CubillasAún no hay calificaciones
- CagadaDocumento24 páginasCagadaQuique Nuevo PerezAún no hay calificaciones
- Cultivo y Composición BioquímicaDocumento12 páginasCultivo y Composición BioquímicaDavidCulquiGarciaAún no hay calificaciones
- BacteriasDocumento3 páginasBacteriasSamantha YañezAún no hay calificaciones
- Pujos Valeria Art 2Documento2 páginasPujos Valeria Art 2Valeria PujosAún no hay calificaciones
- NMP BacteriaHMA UNAMLibroMicrobiologiaDocumento39 páginasNMP BacteriaHMA UNAMLibroMicrobiologiaEliana Sotomayor BohleAún no hay calificaciones
- Descomposición de La Hojarasca y Su Relación Con Los MacroinvertebradosDocumento16 páginasDescomposición de La Hojarasca y Su Relación Con Los MacroinvertebradosAna Belen RobledoAún no hay calificaciones
- Impacto Ecológico de La Industria Petrolera en La Sonda de Campeche, México, Tras Tres Décadas de Actividad - Una RevisiónDocumento16 páginasImpacto Ecológico de La Industria Petrolera en La Sonda de Campeche, México, Tras Tres Décadas de Actividad - Una RevisiónRonaldo Villafuerte AlataAún no hay calificaciones
- Ciencia e Ingeniería Neogranadina 0124-8170: Issn: Revistaing@unimilitar - Edu.coDocumento15 páginasCiencia e Ingeniería Neogranadina 0124-8170: Issn: Revistaing@unimilitar - Edu.cossr.networkmxAún no hay calificaciones
- 2014-Aislamiento y Cultivo de Mi-Croalgas Bentónicas Del Caribe Colombiano Bajo Diferentes Condiciones de TemperaturaDocumento10 páginas2014-Aislamiento y Cultivo de Mi-Croalgas Bentónicas Del Caribe Colombiano Bajo Diferentes Condiciones de Temperaturafaubricio montalvoAún no hay calificaciones
- RMxAA 58-2 - aarellano-VIIIDocumento15 páginasRMxAA 58-2 - aarellano-VIIIStelleAún no hay calificaciones
- The Properties of High Redshift Galaxies: Revista Mexicana de Astronomía y Astrofísica, 58, 83-97 (2022)Documento15 páginasThe Properties of High Redshift Galaxies: Revista Mexicana de Astronomía y Astrofísica, 58, 83-97 (2022)StelleAún no hay calificaciones
- NNP 223 DDocumento3 páginasNNP 223 DStelleAún no hay calificaciones
- RMxAA 58-1 - jliska-XIIDocumento14 páginasRMxAA 58-1 - jliska-XIIStelleAún no hay calificaciones
- RMxAA 58-2 - garreaga-IXDocumento27 páginasRMxAA 58-2 - garreaga-IXStelleAún no hay calificaciones
- RMxAA 58-1 - Ocakirli-IDocumento19 páginasRMxAA 58-1 - Ocakirli-IStelleAún no hay calificaciones
- 54023-Texto Del Artículo-170391-3-10-20230116Documento15 páginas54023-Texto Del Artículo-170391-3-10-20230116StelleAún no hay calificaciones
- 54471-Texto Del Artículo-171316-1-10-20230314Documento9 páginas54471-Texto Del Artículo-171316-1-10-20230314StelleAún no hay calificaciones
- 54386-Texto Del Artículo-171364-1-10-20230316Documento11 páginas54386-Texto Del Artículo-171364-1-10-20230316StelleAún no hay calificaciones
- 54454-Texto Del Artículo-171315-1-10-20230314Documento10 páginas54454-Texto Del Artículo-171315-1-10-20230314StelleAún no hay calificaciones
- NNP 222 ADocumento1 páginaNNP 222 AStelleAún no hay calificaciones
- NNP 223 ADocumento1 páginaNNP 223 AStelleAún no hay calificaciones
- 54577-Texto Del Artículo-169981-1-10-20221026Documento7 páginas54577-Texto Del Artículo-169981-1-10-20221026StelleAún no hay calificaciones
- 1501-Texto Del Artículo-7173-1-10-20230131Documento18 páginas1501-Texto Del Artículo-7173-1-10-20230131StelleAún no hay calificaciones
- 54799-Texto Del Artículo-171299-1-10-20230313Documento14 páginas54799-Texto Del Artículo-171299-1-10-20230313StelleAún no hay calificaciones
- NNP 222 DDocumento14 páginasNNP 222 DStelleAún no hay calificaciones
- 1507-Texto Del Artículo-7210-1-10-20230131Documento22 páginas1507-Texto Del Artículo-7210-1-10-20230131StelleAún no hay calificaciones
- 1508-Texto Del Artículo-7221-1-10-20230131Documento21 páginas1508-Texto Del Artículo-7221-1-10-20230131StelleAún no hay calificaciones
- 1503-Texto Del Artículo-7191-2-10-20230201Documento20 páginas1503-Texto Del Artículo-7191-2-10-20230201StelleAún no hay calificaciones
- NNP 222 CDocumento9 páginasNNP 222 CStelleAún no hay calificaciones
- NNP 222 eDocumento5 páginasNNP 222 eStelleAún no hay calificaciones
- 1506-Texto Del Artículo-7201-1-10-20230131Documento14 páginas1506-Texto Del Artículo-7201-1-10-20230131StelleAún no hay calificaciones
- A200722 GonzalezDocumento14 páginasA200722 GonzalezStelleAún no hay calificaciones
- NNP 223 eDocumento5 páginasNNP 223 eStelleAún no hay calificaciones
- NNP 222 BDocumento7 páginasNNP 222 BStelleAún no hay calificaciones
- NNP 223 CDocumento12 páginasNNP 223 CStelleAún no hay calificaciones
- A180622 DorisonDocumento21 páginasA180622 DorisonStelleAún no hay calificaciones
- A160722 AnjerdiDocumento17 páginasA160722 AnjerdiStelleAún no hay calificaciones
- 1586-Texto Del Artículo-11102-2-10-20221206Documento6 páginas1586-Texto Del Artículo-11102-2-10-20221206StelleAún no hay calificaciones
- A100622 MartinezDocumento21 páginasA100622 MartinezStelleAún no hay calificaciones
- TESIS Tesis Sobre DisfagiaDocumento43 páginasTESIS Tesis Sobre DisfagiaLESLIE GISSEL CAYUPIL DÍAZAún no hay calificaciones
- Preguntas NeumoDocumento41 páginasPreguntas Neumodenisse floresAún no hay calificaciones
- HalitosisDocumento8 páginasHalitosisXime SoledadAún no hay calificaciones
- EjercicioCPT-4Verificando Aprendizaje (1) 56ptsDocumento4 páginasEjercicioCPT-4Verificando Aprendizaje (1) 56ptsalexs_988Aún no hay calificaciones
- Examen General de Orina PoeDocumento4 páginasExamen General de Orina PoeJulieta Gutierrez83% (12)
- TAREA 6 Microbiologia LabDocumento2 páginasTAREA 6 Microbiologia LabRenzo RiveraAún no hay calificaciones
- Exploración de TóraxDocumento14 páginasExploración de TóraxSophie De Arr-van GoghAún no hay calificaciones
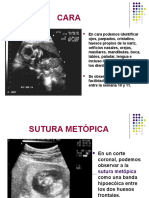
- Tecnicas Ecográficas para Cara, Cuello, Hombro FetalDocumento20 páginasTecnicas Ecográficas para Cara, Cuello, Hombro FetalLaura LizardoAún no hay calificaciones
- Qué Ocurre en El Cerebro de Una Persona Con DislexiaDocumento10 páginasQué Ocurre en El Cerebro de Una Persona Con DislexiaHasnaa MaríaAún no hay calificaciones
- Catatonia, Un Síndrome Nueropsiquiátrico PDFDocumento16 páginasCatatonia, Un Síndrome Nueropsiquiátrico PDFClaudiaPereiraAún no hay calificaciones
- Identificación de Nematodos en Muestras de Suelo Del Cultivo MaracuyáDocumento9 páginasIdentificación de Nematodos en Muestras de Suelo Del Cultivo MaracuyáEDSON MONTENEGRO RODRIGUEZAún no hay calificaciones
- Temario Por Año de Residencia-2Documento9 páginasTemario Por Año de Residencia-2LudmilaRossiAún no hay calificaciones
- Trip Cancer ItzelDocumento2 páginasTrip Cancer ItzelItzel Rubio100% (1)
- HepatectomiaDocumento23 páginasHepatectomiaKyara GuevaraAún no hay calificaciones
- Acupuntura y GinecologiaDocumento4 páginasAcupuntura y GinecologiaAndres GonzalezAún no hay calificaciones
- Silabo Del Curso Semiología ClínicaDocumento9 páginasSilabo Del Curso Semiología ClínicaUsuario1234100% (2)
- XENOBIÓTICODocumento11 páginasXENOBIÓTICOIsabela WilchesAún no hay calificaciones
- Proyecto MacaDocumento4 páginasProyecto MacaJose Eduardo Becerra BazanAún no hay calificaciones
- Tema 1 HemostasiaDocumento127 páginasTema 1 HemostasiaBrigitteMoranAún no hay calificaciones
- Rodriguez Sutil Escision y Disociacion CeIR V9N2Documento18 páginasRodriguez Sutil Escision y Disociacion CeIR V9N2lulopeAún no hay calificaciones
- Oncologia de BolsilloDocumento356 páginasOncologia de Bolsilloalejandra100% (2)
- Tesis: para Optar El Título Profesional De: Licenciada en EnfermeríaDocumento83 páginasTesis: para Optar El Título Profesional De: Licenciada en EnfermeríaCarlos J Flores VacaAún no hay calificaciones
- Hipertensión ArterialDocumento18 páginasHipertensión ArterialAnonymous psFkGlIJ100% (1)
- Concurso de Oposición Docente Pruebas Cognitivas Frecuencia NuméricaDocumento9 páginasConcurso de Oposición Docente Pruebas Cognitivas Frecuencia NuméricaGenesis ConcepcionAún no hay calificaciones
- Laboratorio de Inmunología 2023-1Documento32 páginasLaboratorio de Inmunología 2023-1Enzo PuentesAún no hay calificaciones
- Reporte Materno 2019Documento1 páginaReporte Materno 2019GASTON WALTER FERNANDEZ ESPINOZAAún no hay calificaciones
- Ca. de MamaDocumento10 páginasCa. de MamaCarlos OlveraAún no hay calificaciones
- Memorias Carteles Final15Documento74 páginasMemorias Carteles Final15Carlos CortésAún no hay calificaciones
- Un Ramo de Estructuras de ADNDocumento11 páginasUn Ramo de Estructuras de ADNHector José Ortiz AlvarezAún no hay calificaciones
- Bronquiolitis AgudaaaDocumento5 páginasBronquiolitis AgudaaaALEJANDRO COUTIÑO VALDOVINOSAún no hay calificaciones