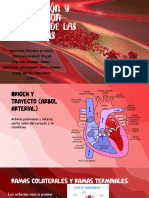
También podría gustarte
- ANATO Preguntero de Final de Anato 2 (2) .Docx Versión 1Documento24 páginasANATO Preguntero de Final de Anato 2 (2) .Docx Versión 1matego41Aún no hay calificaciones
- Examen CASA DE ANATOMIA E HISTOLOGÍA HUMANADocumento25 páginasExamen CASA DE ANATOMIA E HISTOLOGÍA HUMANALinda Fernanda Cruz MorenoAún no hay calificaciones
- Sistema CardiovascularDocumento6 páginasSistema CardiovascularEva PastorAún no hay calificaciones
- CardiovascularDocumento36 páginasCardiovascularDarlene CondoriAún no hay calificaciones
- 02-Cuestionario Cardiovascular 2Documento4 páginas02-Cuestionario Cardiovascular 2Betz KeisukeAún no hay calificaciones
- Clasificación y Descripción Anatómica de Las ArteriasDocumento22 páginasClasificación y Descripción Anatómica de Las ArteriasGómez Cuevas MarcelaAún no hay calificaciones
- Barrera HematoencefalicaDocumento85 páginasBarrera Hematoencefalicaangela jimenezAún no hay calificaciones
- Tejidos de Animales para Cuarto de SecundariaDocumento6 páginasTejidos de Animales para Cuarto de SecundariaClaudia Paola Marina RobalinoAún no hay calificaciones
- CascoDocumento38 páginasCascoCeleste LombardoAún no hay calificaciones
- Lab 7 Cordón UmbilicalDocumento8 páginasLab 7 Cordón Umbilicalkathia jaenAún no hay calificaciones
- Laboratorio Tejido MuscularDocumento8 páginasLaboratorio Tejido MuscularDaniel GarciaAún no hay calificaciones
- Preparados IntegradorDocumento5 páginasPreparados Integradordanyelbatista24Aún no hay calificaciones
- Cuadro Comparativo (Capilares)Documento4 páginasCuadro Comparativo (Capilares)Sun Min-Kyu100% (1)
- Sistema Circulatorio y LinfaticoDocumento9 páginasSistema Circulatorio y LinfaticoSaul MandujanoAún no hay calificaciones
- Lobulillo HepáticoDocumento12 páginasLobulillo Hepáticoyarumi lopez arbaiza0% (1)
- Aparato CirculatorioDocumento8 páginasAparato Circulatorioareli100% (1)
- Tercera UnidadDocumento77 páginasTercera UnidadEricka Ixcamparic PachecoAún no hay calificaciones
- Estructura Microscopica y Funcion Glomerual Renal 4.3.5 SV PDFDocumento36 páginasEstructura Microscopica y Funcion Glomerual Renal 4.3.5 SV PDFCarolina AlitzelAún no hay calificaciones
- Sisema Cardiovascular Aleximar BetancourtDocumento9 páginasSisema Cardiovascular Aleximar BetancourtAdrian BetancourtAún no hay calificaciones
- Resumen Histoembrio Versión 2 Histo .Documento124 páginasResumen Histoembrio Versión 2 Histo .Geo CheAún no hay calificaciones
- ArteriolDocumento2 páginasArteriolFranz Cotache rojasAún no hay calificaciones
- Introduccion A La HIstologiaDocumento102 páginasIntroduccion A La HIstologiaPablo Sanchez MartinezAún no hay calificaciones
- Sistema de Investigacion Venas y OdontologicoDocumento19 páginasSistema de Investigacion Venas y OdontologicoGomez AlejandroAún no hay calificaciones
- Generalidades de CirculatorioDocumento8 páginasGeneralidades de CirculatorioFlorencia Valenzuela Escobar.Aún no hay calificaciones
- Sistema Cardiovascular 2012Documento39 páginasSistema Cardiovascular 2012infodontoulaAún no hay calificaciones
- Practica PVN ADocumento45 páginasPractica PVN AIgnacio BlagodyrAún no hay calificaciones
- Preguntas Fase IiiDocumento9 páginasPreguntas Fase IiiJosé UrdayAún no hay calificaciones
- Organos Del Sistema LinfaticoDocumento14 páginasOrganos Del Sistema LinfaticoMoises GarciaAún no hay calificaciones
- Tema 09,10,11,12Documento15 páginasTema 09,10,11,12Henry LeonardoAún no hay calificaciones
- Histologia 2017 - Aparato Urinario PracticaDocumento64 páginasHistologia 2017 - Aparato Urinario PracticaAnngie Lisbeth Requejo MasAún no hay calificaciones
- Temas Del 1er Parcial. Histologia-1Documento33 páginasTemas Del 1er Parcial. Histologia-1Cristian MorenoAún no hay calificaciones
- Histo Practica Ciclo IiDocumento25 páginasHisto Practica Ciclo IiLeonardo MarrufoAún no hay calificaciones
- Sistema Cardiovascular 2Documento4 páginasSistema Cardiovascular 2rossipe2007Aún no hay calificaciones
- Sistema Circulatorio PDFDocumento5 páginasSistema Circulatorio PDFXimena BonillaAún no hay calificaciones
- Aparato Cardiovascular AumedDocumento36 páginasAparato Cardiovascular AumedPaula LocatelliAún no hay calificaciones
- Cuestionario Sistemas CardiovascularDocumento4 páginasCuestionario Sistemas CardiovascularJahaira CapellanAún no hay calificaciones
- Clase 3 Histologia para MorfologiaDocumento74 páginasClase 3 Histologia para MorfologiaserpenterxAún no hay calificaciones
- Vasos SanguíneosDocumento7 páginasVasos SanguíneosRamiro MadriagaAún no hay calificaciones
- Huesos Del Cuerpo Humano Examen PDFDocumento28 páginasHuesos Del Cuerpo Humano Examen PDFluckapuestasgngioviAún no hay calificaciones
- Lista de Preparados HistologicosDocumento12 páginasLista de Preparados HistologicosSergio Emanuel Santiago DiazAún no hay calificaciones
- Lab CardiovascularDocumento10 páginasLab CardiovascularMarco SossaAún no hay calificaciones
- Juárez Giselle CardiovascularDocumento7 páginasJuárez Giselle CardiovascularGiselle JuarezAún no hay calificaciones
- h8 - Aparato Cardiovascular 2013 53-60 WebDocumento8 páginash8 - Aparato Cardiovascular 2013 53-60 Webjuli2990Aún no hay calificaciones
- LABORATORIO 5 HistoDocumento12 páginasLABORATORIO 5 HistoBrayan F.MAún no hay calificaciones
- Histologia Arterias Venas y CapilaresDocumento45 páginasHistologia Arterias Venas y CapilaresJorge Bautistad100% (3)
- Taller Cuerpo HumanoDocumento11 páginasTaller Cuerpo HumanoStay AcostaAún no hay calificaciones
- Sistema CirculatorioDocumento9 páginasSistema CirculatorioMemo AlvarezAún no hay calificaciones
- Histología Del Aparato CardiovascularDocumento16 páginasHistología Del Aparato CardiovascularstivenAún no hay calificaciones
- Resumen AnatoDocumento6 páginasResumen AnatoCamilaAún no hay calificaciones
- CardiovascularDocumento58 páginasCardiovascularMAURICIO ALEXANDER LIMACHE MERMAAún no hay calificaciones
- Tarea 3ra Unidad: Alumna: Ericka Maria Alejandra Ixcamparic Pacheco Sección: ADocumento26 páginasTarea 3ra Unidad: Alumna: Ericka Maria Alejandra Ixcamparic Pacheco Sección: AEricka Ixcamparic PachecoAún no hay calificaciones
- PRACTICA III de HistologiaDocumento60 páginasPRACTICA III de HistologiaKaren Grajeda RuizAún no hay calificaciones
- Informe Sesion 02Documento13 páginasInforme Sesion 02NAYELLY JAMILETH LOPEZ PAREDESAún no hay calificaciones
- SISTEMA CARDIOVASCULAR-Parte 2Documento44 páginasSISTEMA CARDIOVASCULAR-Parte 2Alexandra Escontrias LaraAún no hay calificaciones
- Tejido EpitelialDocumento5 páginasTejido EpitelialnorkyAún no hay calificaciones
- Histologia Presentación Corazon MiaDocumento60 páginasHistologia Presentación Corazon MiaKarina BarbosaAún no hay calificaciones
- Sistema Reproductor Masculino y FemeninoDocumento17 páginasSistema Reproductor Masculino y FemeninoDulce RosalesAún no hay calificaciones
- Tejidos de Animales para Cuarto de SecundariaDocumento6 páginasTejidos de Animales para Cuarto de Secundariawiliam Calderon FloresAún no hay calificaciones
- La CirculaciónDocumento4 páginasLa CirculaciónDíaz Zumaeta Ariana MichelleAún no hay calificaciones
- Cuestionario 8Documento1 páginaCuestionario 8Chris KeoAún no hay calificaciones
- Actividad 4 de La 10Documento5 páginasActividad 4 de La 10Chris KeoAún no hay calificaciones
- BioquimicaDocumento3 páginasBioquimicaChris KeoAún no hay calificaciones
- Examen 2Documento2 páginasExamen 2Chris KeoAún no hay calificaciones
- Perfil AlcantarilladoDocumento1 páginaPerfil AlcantarilladoDiego Ortiz MolinaAún no hay calificaciones
- REFUERZO GRADO 11° Ciencias NaturalesDocumento5 páginasREFUERZO GRADO 11° Ciencias Naturalesmiressa rangel0% (1)
- Guía Practica - Embriogenesis - Grupo 1 - NRC4038Documento7 páginasGuía Practica - Embriogenesis - Grupo 1 - NRC4038MabelAún no hay calificaciones
- Nuestra Tierra 00Documento23 páginasNuestra Tierra 00nicolasAún no hay calificaciones
- Clase Virtual - FotosintesisDocumento8 páginasClase Virtual - FotosintesisAlexandra FloresAún no hay calificaciones
- Desempeños para Estudiantes Con DiscapacidadDocumento6 páginasDesempeños para Estudiantes Con Discapacidadjavier orlando montenegro garzonAún no hay calificaciones
- Mecanismo Fisiológico de Filtración Del Riñón y Cómo Se Relaciona Con La Presión ArterialDocumento5 páginasMecanismo Fisiológico de Filtración Del Riñón y Cómo Se Relaciona Con La Presión ArterialLarissa Aban.Aún no hay calificaciones
- CADERADocumento6 páginasCADERAf6zvj652rkAún no hay calificaciones
- Establecimiento in Vitro de Yemas de PassifloraDocumento4 páginasEstablecimiento in Vitro de Yemas de Passifloraalma ramirezAún no hay calificaciones
- Mecanismos de Envejecimiento CelularDocumento8 páginasMecanismos de Envejecimiento CelularTgbAún no hay calificaciones
- Hoja de Registro en Vias de Pavimento Flexible: Doble Vía Rio Seco-Huarina Izquierda 8+780 01 08 2/21/2019 8+820 7.00 m2Documento10 páginasHoja de Registro en Vias de Pavimento Flexible: Doble Vía Rio Seco-Huarina Izquierda 8+780 01 08 2/21/2019 8+820 7.00 m2jorge perezAún no hay calificaciones
- Gráfico de Mapa Mental o Conceptual de Seis Ideas Relacionadas A Una Idea Principal Con Formas Dibujado A Mano MulticolorDocumento1 páginaGráfico de Mapa Mental o Conceptual de Seis Ideas Relacionadas A Una Idea Principal Con Formas Dibujado A Mano Multicolorandy paul villanueva gironAún no hay calificaciones
- Un Curso de FelicidadDocumento157 páginasUn Curso de FelicidadJose David Cohen Rhenals100% (2)
- Eliminacion de OracionesDocumento2 páginasEliminacion de OracioneszimoneAún no hay calificaciones
- Guia Tema 01 Morfofisiologia BucalDocumento20 páginasGuia Tema 01 Morfofisiologia BucalpabloAún no hay calificaciones
- Anatomía y Fisiología RenalDocumento7 páginasAnatomía y Fisiología RenalFrancisco MontañoAún no hay calificaciones
- Plandemanejo Uac LlasDocumento149 páginasPlandemanejo Uac LlasJhonatan Andres Arias ZapataAún no hay calificaciones
- Informe de La CelulaDocumento16 páginasInforme de La Celulareisly almonteAún no hay calificaciones
- 51 Consejos para Vivir Una Vida MejorDocumento18 páginas51 Consejos para Vivir Una Vida MejorMax PortAún no hay calificaciones
- Celulas Eucariotas y Procariotas-Sem.3Documento32 páginasCelulas Eucariotas y Procariotas-Sem.3Manuel CespedesAún no hay calificaciones
- Sistemas Ascendentes Sensoriales y Procesamiento Sensorial CentralDocumento5 páginasSistemas Ascendentes Sensoriales y Procesamiento Sensorial CentralFhernye AlbaAún no hay calificaciones
- Aparato Digestivo de Algunos Invertebrados y Algunos TemasDocumento7 páginasAparato Digestivo de Algunos Invertebrados y Algunos TemasJESUS TURPO PAUCARAún no hay calificaciones
- OSIFICACIÓNDocumento4 páginasOSIFICACIÓNNIKCOL LORENA JARAMILLO ACOSTAAún no hay calificaciones
- Tesis ArgumentativoDocumento7 páginasTesis Argumentativosantiago lopez valenciaAún no hay calificaciones
- Preguntas Sobre Cavidades CorporalesDocumento2 páginasPreguntas Sobre Cavidades CorporalesJorge Andres LozanoAún no hay calificaciones
- Tarea 7 CriminalisticaDocumento5 páginasTarea 7 CriminalisticaDominga PolancoAún no hay calificaciones
- El Cuerpo Humano - Secuencia DidacticaDocumento116 páginasEl Cuerpo Humano - Secuencia DidacticaTamara SeñoAún no hay calificaciones
- Adn de Tomate PDFDocumento7 páginasAdn de Tomate PDFFighterAún no hay calificaciones
- Clase 2Documento11 páginasClase 2BRAYAN LEONEL GUTIERREZ CHACONAún no hay calificaciones
- Ecologia AceptadaDocumento12 páginasEcologia AceptadaSergio Andrés PrietoAún no hay calificaciones