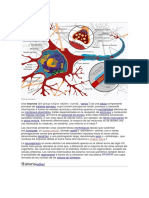
También podría gustarte
- NeuronaDocumento19 páginasNeuronaYury Cruz VillegasAún no hay calificaciones
- Neurona: unidad estructural y funcional del sistema nerviosoDocumento9 páginasNeurona: unidad estructural y funcional del sistema nerviosomarthaAún no hay calificaciones
- NeuronaDocumento24 páginasNeuronaAlejandra BonillaAún no hay calificaciones
- Sistema Nervioso Central, Periferico y NeuronaDocumento28 páginasSistema Nervioso Central, Periferico y NeuronasalvayopsAún no hay calificaciones
- Neuron ADocumento11 páginasNeuron Aalexandra leivaAún no hay calificaciones
- NeuronasDocumento6 páginasNeuronasmiriam vargas holguinAún no hay calificaciones
- NeuronaDocumento2 páginasNeuronaRodolfo LatorreAún no hay calificaciones
- Las NeuronasDocumento13 páginasLas NeuronasGladys Hernández JiménezAún no hay calificaciones
- Copia de Libro de TRASTORNO DE NEURODESARROLLODocumento351 páginasCopia de Libro de TRASTORNO DE NEURODESARROLLOnathalyrendonAún no hay calificaciones
- Leira 7 PDFDocumento7 páginasLeira 7 PDFPatricia PorrinoAún no hay calificaciones
- NeuronaDocumento59 páginasNeuronahfuentes0100% (1)
- 5991-BT-02-18-Fisiologà A Neuronal - Potencial Accià N e Impulso Nervioso - 7%Documento20 páginas5991-BT-02-18-Fisiologà A Neuronal - Potencial Accià N e Impulso Nervioso - 7%Luz Tapia VillarroelAún no hay calificaciones
- Células del Sistema Nervioso: Historia y TécnicasDocumento29 páginasCélulas del Sistema Nervioso: Historia y TécnicasLorei DaAún no hay calificaciones
- Leira 7 PDFDocumento7 páginasLeira 7 PDFGiu PaoAún no hay calificaciones
- Guía de Aprendizaje Las Células NeuronalesDocumento7 páginasGuía de Aprendizaje Las Células NeuronalesCelinda GuzmanAún no hay calificaciones
- Las Células Del Sistema NerviosoDocumento16 páginasLas Células Del Sistema Nerviosog.colleenAún no hay calificaciones
- Neuron ADocumento5 páginasNeuron Amax_14_15Aún no hay calificaciones
- Neurona TrabajoDocumento9 páginasNeurona TrabajoMalvina Soledad TorresAún no hay calificaciones
- AnatonullDocumento24 páginasAnatonullRaysa OrtizAún no hay calificaciones
- Una Neurona Típica Consta deDocumento5 páginasUna Neurona Típica Consta deMargarita Piña RodriguezAún no hay calificaciones
- La neurona como unidad funcional del SNCDocumento33 páginasLa neurona como unidad funcional del SNCMaximiliano LozaAún no hay calificaciones
- El arco reflejo y sus partesDocumento27 páginasEl arco reflejo y sus partesMarielba Velandia Lozada100% (1)
- NeuronaDocumento8 páginasNeuronaReyna RodriguezAún no hay calificaciones
- Guia Biologia °10 Las NeuronasDocumento9 páginasGuia Biologia °10 Las NeuronasSialca GestiónAún no hay calificaciones
- Neurona estructura función 40Documento15 páginasNeurona estructura función 40Nico Ignacio VeraAún no hay calificaciones
- NeuronaDocumento2 páginasNeuronaMaferGonzalezAún no hay calificaciones
- La NeuronaDocumento14 páginasLa NeuronaElizabeth Akary Rizo Nishikido100% (2)
- Tema 2. La NeuronaDocumento40 páginasTema 2. La NeuronaIsai JuniorAún no hay calificaciones
- Anatomía de La NeuronaDocumento7 páginasAnatomía de La NeuronaClaudita G. Domínguez PradoAún no hay calificaciones
- Neuroanatomía UFLPDocumento61 páginasNeuroanatomía UFLPluis navarro salgadoAún no hay calificaciones
- Guía de Aprendizaje NeuronaDocumento4 páginasGuía de Aprendizaje Neuronatercerom2009100% (2)
- Las Neuronas y Su HistoriaDocumento3 páginasLas Neuronas y Su HistoriawidmyAún no hay calificaciones
- Tarea SNCDocumento10 páginasTarea SNCAmmy PazmiñoAún no hay calificaciones
- Histología del Sistema Nervioso - U2 _ Parte 4 (1)Documento3 páginasHistología del Sistema Nervioso - U2 _ Parte 4 (1)Daniela Figueroa de RayonAún no hay calificaciones
- GlosarioDocumento10 páginasGlosariokatty posliguaAún no hay calificaciones
- 01 La NeuronaDocumento36 páginas01 La NeuronaStephany DessiréAún no hay calificaciones
- Resumen NeurocienciasDocumento9 páginasResumen NeurocienciasbriAún no hay calificaciones
- Fisiología del Sistema Nervioso Central en NeuropsicologíaDocumento22 páginasFisiología del Sistema Nervioso Central en NeuropsicologíaAna Paula GarciaAún no hay calificaciones
- Tipos Cel Neurona PDFDocumento13 páginasTipos Cel Neurona PDFGracias DiosAún no hay calificaciones
- Sistema NerviosoDocumento11 páginasSistema NerviosoSaúl Ramírez IñigoAún no hay calificaciones
- Tarea 7.1 Tejido Nervioso HISTO 1Documento7 páginasTarea 7.1 Tejido Nervioso HISTO 1Elizabeth EstevezAún no hay calificaciones
- Neurociencia Aplicada 2007 PDFDocumento30 páginasNeurociencia Aplicada 2007 PDFBrayan CorderoAún no hay calificaciones
- Tejido Nervioso AnimalDocumento27 páginasTejido Nervioso AnimalCarmen Luzmila Carlos de la CruzAún no hay calificaciones
- Histología Del Sistema Nervioso CentralDocumento25 páginasHistología Del Sistema Nervioso CentralCesar Garcia NKAún no hay calificaciones
- Fisiologia - Sistema - Nervioso - PPT ExpoDocumento45 páginasFisiologia - Sistema - Nervioso - PPT ExpoSalud Medicina Confianza0% (1)
- Tejido Nervioso ApunteDocumento39 páginasTejido Nervioso ApunteFacu CobosAún no hay calificaciones
- Tejido NerviosoDocumento41 páginasTejido NerviosoItaty JaramilloAún no hay calificaciones
- Tarea 2 La NeuronaDocumento19 páginasTarea 2 La NeuronaClaudia de la CruzAún no hay calificaciones
- Introduccion Basica VV 0.202110 PDFDocumento19 páginasIntroduccion Basica VV 0.202110 PDFJeannette del RosarioAún no hay calificaciones
- Tema 4 Fisiologia DEFINITIVODocumento28 páginasTema 4 Fisiologia DEFINITIVOmagalyamazonasaAún no hay calificaciones
- El Tejido NerviosoDocumento3 páginasEl Tejido NerviosoRafael LinaresAún no hay calificaciones
- Células GlialesDocumento4 páginasCélulas GlialesMaria Reyna Guerrero BalderasAún no hay calificaciones
- 10 - Udd 1A - BiologiaDocumento6 páginas10 - Udd 1A - BiologiaAndrea ChavarriaAún no hay calificaciones
- Actividad 1 NeuronaDocumento3 páginasActividad 1 Neuronatatiane camposAún no hay calificaciones
- La Estructura y La Función de La NeuronaDocumento17 páginasLa Estructura y La Función de La NeuronaChristhopher Miguel Copa Mamani100% (1)
- Introducción A La Neurociencia - BrainwareDocumento51 páginasIntroducción A La Neurociencia - Brainwaremateo pianaAún no hay calificaciones
- Anatomia y Fisiología del Sistema Nervioso IIDe EverandAnatomia y Fisiología del Sistema Nervioso IICalificación: 4 de 5 estrellas4/5 (28)
- Neuropsicología, Conceptos y TecnicasDocumento26 páginasNeuropsicología, Conceptos y Tecnicasangel esteban palomino bedoyaAún no hay calificaciones
- Resumen Neurociencia Tema 1Documento8 páginasResumen Neurociencia Tema 1Nayara Solano JiménezAún no hay calificaciones
- AstrocitosDocumento3 páginasAstrocitosJake SolivanAún no hay calificaciones
- CarcinomaDocumento6 páginasCarcinomaJake SolivanAún no hay calificaciones
- DiabetesDocumento2 páginasDiabetesJake SolivanAún no hay calificaciones
- MeconeoDocumento1 páginaMeconeoJake SolivanAún no hay calificaciones
- Evolución homínidos ÁfricaDocumento3 páginasEvolución homínidos ÁfricaTamara MuñozAún no hay calificaciones
- Libro RV CompletoDocumento306 páginasLibro RV CompletoJose Maria Martinelli CostaAún no hay calificaciones
- Practica 2Documento10 páginasPractica 2Diana Sanchez sanchezAún no hay calificaciones
- Estudio microbiología oral cavidad bocaDocumento18 páginasEstudio microbiología oral cavidad bocaBPMAún no hay calificaciones
- Los seres vivos necesitan agua, aire y alimentoDocumento13 páginasLos seres vivos necesitan agua, aire y alimentoCARMEN ADRIANA CAMACHO SEGURAAún no hay calificaciones
- CORDIS - Article - 24410 British Scientists Create Human Embryos Without Sperm - EsDocumento3 páginasCORDIS - Article - 24410 British Scientists Create Human Embryos Without Sperm - EsAngel DavidAún no hay calificaciones
- Ciencias Naturales ResumenDocumento2 páginasCiencias Naturales ResumenCarlaAún no hay calificaciones
- Grupo 2Documento26 páginasGrupo 2JORGE OLVERAAún no hay calificaciones
- Monografia NEURO CIENCIA PDFDocumento19 páginasMonografia NEURO CIENCIA PDFSheila LopezAún no hay calificaciones
- Bomba de Sodio Potasio - 13Documento6 páginasBomba de Sodio Potasio - 13Ninoska MartinezAún no hay calificaciones
- Tórax o Caja TorácicaDocumento3 páginasTórax o Caja TorácicaAndrea BarzolaAún no hay calificaciones
- 1.4 HomeostasisDocumento10 páginas1.4 HomeostasisMayela StnnAún no hay calificaciones
- AnatomiaDocumento13 páginasAnatomiaUnai 248Aún no hay calificaciones
- Purificación de Una ProteínaDocumento8 páginasPurificación de Una ProteínaasesinamortalAún no hay calificaciones
- ORTHOPTERA NEUROPTERA MastispidaeDocumento9 páginasORTHOPTERA NEUROPTERA Mastispidaedelvis sanchez diasAún no hay calificaciones
- Colecta y Montaje de InsectosDocumento38 páginasColecta y Montaje de InsectosPaul Palacin Guerra100% (2)
- APUNTESDocumento45 páginasAPUNTESSara Rodriguez MolinaAún no hay calificaciones
- ¡Activa Tus Neuronas Ambientales!Documento2 páginas¡Activa Tus Neuronas Ambientales!carolinaruelasarmentaAún no hay calificaciones
- Cuadro SinópticoDocumento2 páginasCuadro SinópticoDaniela InguAún no hay calificaciones
- Analisis Tecnico Propiedad San JoaquinDocumento9 páginasAnalisis Tecnico Propiedad San JoaquinSergio_thaineAún no hay calificaciones
- 10 Canciones GuatemaltecasDocumento13 páginas10 Canciones GuatemaltecasPistacheAún no hay calificaciones
- Bimestral Naturales Grado Sexto 1 PeriodoDocumento4 páginasBimestral Naturales Grado Sexto 1 PeriodoAndres Mauricio Moncada Garcia100% (1)
- Aprendizaje, Coevolución Neuroambiental: Coordinación Educativa Y Cultural CentroamericanaDocumento20 páginasAprendizaje, Coevolución Neuroambiental: Coordinación Educativa Y Cultural CentroamericanaBenny NeiraAún no hay calificaciones
- Sistema TegumentarioDocumento3 páginasSistema TegumentarioGregorio Cortés Emma GuadalupeAún no hay calificaciones
- TP15 GeneticaDocumento9 páginasTP15 GeneticaGab RuecknerAún no hay calificaciones
- 4 Histologia de La Mucosa Bucal 2020 Ciremo PDFDocumento21 páginas4 Histologia de La Mucosa Bucal 2020 Ciremo PDFjosueAún no hay calificaciones
- Aves en Tigre GrandeDocumento89 páginasAves en Tigre GrandeJulia Huchin CocomAún no hay calificaciones
- Nuevos Descubrimientos en La MicrobiologíaDocumento5 páginasNuevos Descubrimientos en La MicrobiologíaFatima Anahy Nuñez MitreAún no hay calificaciones
- Scheler El Puesto Del Hombre en El Cosmos. IntroducciónDocumento4 páginasScheler El Puesto Del Hombre en El Cosmos. IntroducciónNestor SotomayorAún no hay calificaciones
- Folleto de Anatofisiologia Del Corazon Por Carlos Andres Garcia 72140 Downloable 982082Documento37 páginasFolleto de Anatofisiologia Del Corazon Por Carlos Andres Garcia 72140 Downloable 982082Gisela18Aún no hay calificaciones