También podría gustarte
- INFORMEDocumento10 páginasINFORMEluz pari ciertoAún no hay calificaciones
- Competencia LV 17-03-09Documento8 páginasCompetencia LV 17-03-09NaybethGuzmánAún no hay calificaciones
- Ecologia Relaciones InterespecificasDocumento14 páginasEcologia Relaciones Interespecificasjeysstefanny100% (1)
- CLASE 8 Comunidades IDocumento25 páginasCLASE 8 Comunidades Icatalina gerkenAún no hay calificaciones
- Tema 14 Comp Interespecifica 2020Documento27 páginasTema 14 Comp Interespecifica 2020Mario Álvarez GarcíaAún no hay calificaciones
- InteraccionesDocumento30 páginasInteraccionesNataly LunaAún no hay calificaciones
- Luis SenaDocumento5 páginasLuis SenaLuis HdysusksggxAún no hay calificaciones
- EcologíaDocumento15 páginasEcologíaKarianny TorresAún no hay calificaciones
- Practicas Competencia DepredacionDocumento6 páginasPracticas Competencia DepredacionJOSE FRANCISCO VILLASANTE BENAVIDESAún no hay calificaciones
- Taller Modelos CompletoDocumento22 páginasTaller Modelos CompletocarolinaAún no hay calificaciones
- El Modelo DepredadorDocumento4 páginasEl Modelo DepredadorJoy BelenAún no hay calificaciones
- Competencia y ExplotaciónDocumento47 páginasCompetencia y ExplotaciónNapoOlaveAún no hay calificaciones
- Informe 9 Computacional 1Documento3 páginasInforme 9 Computacional 1Luis Carlosviza PuentesAún no hay calificaciones
- Modelos EcologicosDocumento9 páginasModelos EcologicosGermain DE LA CruzAún no hay calificaciones
- Taller 7Documento8 páginasTaller 7Ana Natalia Lopez VargasAún no hay calificaciones
- Todo BioDocumento12 páginasTodo Bionecajas389Aún no hay calificaciones
- Simulación en PopulusDocumento21 páginasSimulación en PopulusMayby Tovar100% (1)
- Taller 7Documento3 páginasTaller 7Catherinee Moora100% (1)
- t9 Sistema Depredador PresaDocumento10 páginast9 Sistema Depredador PresaMario JrrAún no hay calificaciones
- Depredador PresaDocumento9 páginasDepredador PresaJose Alejandro Vargas CordovaAún no hay calificaciones
- Modelo Depredador PresaDocumento32 páginasModelo Depredador PresaEmerson Maucaylle palominoAún no hay calificaciones
- Modelo Depredador PresaDocumento4 páginasModelo Depredador Presamike1521Aún no hay calificaciones
- 3 - Interacciones Entre EspeciesDocumento15 páginas3 - Interacciones Entre EspeciespedroAún no hay calificaciones
- ABA1312Documento2 páginasABA1312David el notasAún no hay calificaciones
- Desarrollo Del Foro 3.Documento9 páginasDesarrollo Del Foro 3.Leo MejiaAún no hay calificaciones
- Ecuaciones Lotka-VolterraDocumento2 páginasEcuaciones Lotka-VolterraLuis MalaverAún no hay calificaciones
- Quítate Tu para Ponerme YoDocumento12 páginasQuítate Tu para Ponerme YoEvelin Castillo Sanchez100% (1)
- Efect o AlleeDocumento17 páginasEfect o AlleeGabriel Crispin SilvaAún no hay calificaciones
- Diversidad Alfa 1206429401560291 3Documento50 páginasDiversidad Alfa 1206429401560291 3Cintia ZalazarAún no hay calificaciones
- Estructura de ComunidadesDocumento18 páginasEstructura de ComunidadesPaula Báez SolanoAún no hay calificaciones
- Inter Acci OnesDocumento40 páginasInter Acci OnesYadira RamírezAún no hay calificaciones
- Proyecto Final MATwordDocumento10 páginasProyecto Final MATwordCarla RamirezAún no hay calificaciones
- Corre Que Te AlcanzoDocumento22 páginasCorre Que Te AlcanzoEvelin Castillo SanchezAún no hay calificaciones
- ComInter 17 5 16 - CompletarDocumento32 páginasComInter 17 5 16 - CompletarmarroquinantoniofaviorobertoAún no hay calificaciones
- Expo Como Varia La Abundancia Relativa de Las EspeciesDocumento8 páginasExpo Como Varia La Abundancia Relativa de Las EspeciesNatalia BravoAún no hay calificaciones
- Apuntes Sobre EcologiaDocumento62 páginasApuntes Sobre EcologiaCentoloAún no hay calificaciones
- 13-Interacciones InterespecíficasDocumento3 páginas13-Interacciones InterespecíficasAmparo PratticoAún no hay calificaciones
- Crecimiento ExponencialDocumento7 páginasCrecimiento ExponencialStefania Janeth Rivera CantilloAún no hay calificaciones
- Depredador PresaDocumento6 páginasDepredador PresaAdriana Lucía Gualy VargasAún no hay calificaciones
- Valvasori Marroncle, Dulce M. - Actividad L-V (EcoPob)Documento2 páginasValvasori Marroncle, Dulce M. - Actividad L-V (EcoPob)Duly ValvasoriAún no hay calificaciones
- t9 Sistema Depredador PresaDocumento10 páginast9 Sistema Depredador PresaQuinio Bruno Espiritu EugenioAún no hay calificaciones
- Empleo de Modelos Matemáticos en EcologíaDocumento8 páginasEmpleo de Modelos Matemáticos en EcologíaEl Perro PotaxioAún no hay calificaciones
- Los Ecólogos Clasifican Las Interacciones Entre Las Especies Según Los Resultados PositivosDocumento20 páginasLos Ecólogos Clasifican Las Interacciones Entre Las Especies Según Los Resultados PositivosAdriana Lucía Gualy VargasAún no hay calificaciones
- Formato - Caso de EstudioDocumento14 páginasFormato - Caso de EstudioLUIS FERNANDO RODRIGUEZ ANCHUNDIAAún no hay calificaciones
- S02 - S03 - PPT2 - Ecología y EcosistemasDocumento61 páginasS02 - S03 - PPT2 - Ecología y EcosistemasFERNANDO DE LA PUENTEAún no hay calificaciones
- Interacciones BiologicasDocumento4 páginasInteracciones BiologicasMedeus MoriartyAún no hay calificaciones
- Biotic AsDocumento1 páginaBiotic AsLUIS_MI52Aún no hay calificaciones
- Angelita HHHHHHDocumento9 páginasAngelita HHHHHHAngela SantanaAún no hay calificaciones
- EcuaDocumento7 páginasEcuaEdwardInfanteAún no hay calificaciones
- Interacción o Interrelación de EspeciesDocumento12 páginasInteracción o Interrelación de EspeciesLau OtteroAún no hay calificaciones
- 3) La Estructura Ausente, Capítulo 2Documento7 páginas3) La Estructura Ausente, Capítulo 2Facundo LoliscioAún no hay calificaciones
- Formato - Caso de EstudioDocumento13 páginasFormato - Caso de EstudioLUIS FERNANDO RODRIGUEZ ANCHUNDIAAún no hay calificaciones
- TERRITORIO Clase 8 PowerPointDocumento22 páginasTERRITORIO Clase 8 PowerPointLizbe Peña MoranAún no hay calificaciones
- Proyecto Cientifico 6-Semana 2 OctavoDocumento7 páginasProyecto Cientifico 6-Semana 2 OctavoGabby MoreiraAún no hay calificaciones
- Presa DepredadorDocumento11 páginasPresa DepredadorAldo JuarezAún no hay calificaciones
- Lotka VolterraDocumento10 páginasLotka VolterraYersi Salvador PorrasAún no hay calificaciones
- TP 10 DiversidadDocumento8 páginasTP 10 DiversidadmicaelaAún no hay calificaciones
- FECONIDocumento19 páginasFECONIAdriana CarranzaAún no hay calificaciones
- DermatologiaDocumento3 páginasDermatologiaRukkalaf VeterinariaAún no hay calificaciones
- Centro RespiratórioDocumento4 páginasCentro RespiratórioVitória SeverinoAún no hay calificaciones
- Plan de Mejoramiento Ciencias 5 Periodo 2Documento8 páginasPlan de Mejoramiento Ciencias 5 Periodo 2ELIZABETH BRICEÑOAún no hay calificaciones
- El Pool de AminoácidosDocumento9 páginasEl Pool de AminoácidosKarla Torres100% (1)
- Lectura. Animales en Peligro de ExtinciónDocumento5 páginasLectura. Animales en Peligro de ExtinciónELIZABETHAún no hay calificaciones
- Orden HymenopteraDocumento23 páginasOrden HymenopteraJHON STEVEEN CUSCUE SILVAAún no hay calificaciones
- ArqueobacteriasDocumento2 páginasArqueobacteriaspedro meyer alvaron cristobalAún no hay calificaciones
- Atemi 10 WGDocumento10 páginasAtemi 10 WGCarlos Armando MaldonadoAún no hay calificaciones
- Poder de La Mente2000 RadionicaDocumento317 páginasPoder de La Mente2000 RadionicaCarlos Zelaya100% (5)
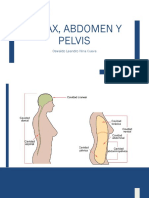
- 5 Tórax, Abdomen y PelvisDocumento195 páginas5 Tórax, Abdomen y PelvisMariecieloLeonAún no hay calificaciones
- Propedeutica ObstetriciaDocumento51 páginasPropedeutica ObstetriciaFernando Paz100% (5)
- Guía Módulo III Escuela Del AguaDocumento122 páginasGuía Módulo III Escuela Del AguaYessica Ruiz100% (1)
- Cuadro Sinoptico - FUNCIONES BIOMECANICAS DEL APARATO MOTORDocumento2 páginasCuadro Sinoptico - FUNCIONES BIOMECANICAS DEL APARATO MOTORAnii ChavezAún no hay calificaciones
- Abonos Verdes y PraderasDocumento5 páginasAbonos Verdes y PraderaspabloAún no hay calificaciones
- Proyecto Farmaceutico ITGAM 2Documento18 páginasProyecto Farmaceutico ITGAM 2Jesús Demetrio Ramírez MoralesAún no hay calificaciones
- 7 M Complementario Sobre ReplicaciónDocumento5 páginas7 M Complementario Sobre ReplicaciónLudmila Aquino BiscaldiAún no hay calificaciones
- Dieta Sin Gluten y Sin CaseinaDocumento6 páginasDieta Sin Gluten y Sin CaseinaAprendizaje Significativo CiberbbAún no hay calificaciones
- Insuficiencia SuprarrenalDocumento13 páginasInsuficiencia SuprarrenalYesenia Indira Montoya100% (1)
- Perspectivas de La Biodiversidad en Centroamerica 2003Documento108 páginasPerspectivas de La Biodiversidad en Centroamerica 2003pips rocAún no hay calificaciones
- AA4 Evidencia Diseno de Instrumentos EvaluativoDocumento11 páginasAA4 Evidencia Diseno de Instrumentos Evaluativojorge carrilloAún no hay calificaciones
- Banco Anato ExitosDocumento49 páginasBanco Anato ExitosWilhelm SánchezAún no hay calificaciones
- Mejoramiento Geneticos en PlantasDocumento36 páginasMejoramiento Geneticos en PlantascastcarlAún no hay calificaciones
- Libro Musica PDFDocumento256 páginasLibro Musica PDFsdppgAún no hay calificaciones
- LípidoDocumento15 páginasLípidogerado033Aún no hay calificaciones
- Daños Ocasionados Por MalezasDocumento12 páginasDaños Ocasionados Por Malezaszamorano3Aún no hay calificaciones
- 61 Comunicacion AsertivaDocumento4 páginas61 Comunicacion AsertivaMarcos Gutierrez Aquino100% (1)
- PC1 Fila A - Mec. - de - MaterialesDocumento3 páginasPC1 Fila A - Mec. - de - MaterialesAngel Esteban Palomino BenitesAún no hay calificaciones
- Corallina (Corallina Elongata) - v3Documento2 páginasCorallina (Corallina Elongata) - v3Aracelli Coayla MurilloAún no hay calificaciones
- GTP T5.células 3 Parte Membrana Plasmática 2015-17Documento54 páginasGTP T5.células 3 Parte Membrana Plasmática 2015-17Maria AraujoAún no hay calificaciones
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- Historia del cuerpo humano: Evolución, salud y enfermedadDe EverandHistoria del cuerpo humano: Evolución, salud y enfermedadAún no hay calificaciones
- Sesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesDe EverandSesgos Cognitivos: Una Fascinante Mirada dentro de la Psicología Humana y los Métodos para Evitar la Disonancia Cognitiva, Mejorar sus Habilidades para Resolver Problemas y Tomar Mejores DecisionesCalificación: 4.5 de 5 estrellas4.5/5 (13)
- Homo antecessor: El nacimiento de una especieDe EverandHomo antecessor: El nacimiento de una especieCalificación: 5 de 5 estrellas5/5 (1)
- Cuerpo Tóxico: Como Liberar Tu Cuerpo De Las Toxinas Externas E Internas, Y Evitar Asi Los Efectos De Los Radicales LibresDe EverandCuerpo Tóxico: Como Liberar Tu Cuerpo De Las Toxinas Externas E Internas, Y Evitar Asi Los Efectos De Los Radicales LibresCalificación: 5 de 5 estrellas5/5 (2)
- 50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaDe Everand50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaCalificación: 4 de 5 estrellas4/5 (37)
- Diseño, construcción y pruebas de horno prototipo semicontinuo para producir cerámicaDe EverandDiseño, construcción y pruebas de horno prototipo semicontinuo para producir cerámicaAún no hay calificaciones
- Liberación del trauma: Perdón y temblor es el caminoDe EverandLiberación del trauma: Perdón y temblor es el caminoCalificación: 4 de 5 estrellas4/5 (4)
- Anatomía del caballo: Guía práctica ilustradaDe EverandAnatomía del caballo: Guía práctica ilustradaCalificación: 4 de 5 estrellas4/5 (9)
- GuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisDe EverandGuíaBurros Análisis clínicos: Todo lo que necesitas saber para entender tus análisisCalificación: 4 de 5 estrellas4/5 (9)
- Inteligencia artificial: Una exploración filosófica sobre el futuro de la mente y la concienciaDe EverandInteligencia artificial: Una exploración filosófica sobre el futuro de la mente y la concienciaAna Isabel Sánchez DíezCalificación: 4 de 5 estrellas4/5 (3)
- Cerebro y silencio: Las claves de la creatividad y la serenidadDe EverandCerebro y silencio: Las claves de la creatividad y la serenidadCalificación: 5 de 5 estrellas5/5 (2)
- La increíble conexión intestino cerebro: Descubre la relación entre las emociones y el equilibrio intestinalDe EverandLa increíble conexión intestino cerebro: Descubre la relación entre las emociones y el equilibrio intestinalCalificación: 4.5 de 5 estrellas4.5/5 (103)
- Cerebros rotos: Pacientes asombrosos que me enseñaron a vivirDe EverandCerebros rotos: Pacientes asombrosos que me enseñaron a vivirCalificación: 5 de 5 estrellas5/5 (3)
- Psicología forense: Estudio de la mente criminalDe EverandPsicología forense: Estudio de la mente criminalCalificación: 4.5 de 5 estrellas4.5/5 (18)
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- Genética general: Libro de textoDe EverandGenética general: Libro de textoCalificación: 4.5 de 5 estrellas4.5/5 (11)
- UF0085 - Diagnóstico estético facial y corporalDe EverandUF0085 - Diagnóstico estético facial y corporalCalificación: 4 de 5 estrellas4/5 (5)
- El nacimiento del pensamiento científico: Anaximandro de MiletoDe EverandEl nacimiento del pensamiento científico: Anaximandro de MiletoCalificación: 4.5 de 5 estrellas4.5/5 (4)
- 200 tareas en terapia breve: 2ª ediciónDe Everand200 tareas en terapia breve: 2ª ediciónCalificación: 4.5 de 5 estrellas4.5/5 (33)
- Manual de osteología de cráneo bovino y equinoDe EverandManual de osteología de cráneo bovino y equinoCalificación: 4.5 de 5 estrellas4.5/5 (7)
- El nervio vago. Su poder sanador: Técnicas para tratar la depresión, la ansiedad, los traumas y otros problemasDe EverandEl nervio vago. Su poder sanador: Técnicas para tratar la depresión, la ansiedad, los traumas y otros problemasCalificación: 4.5 de 5 estrellas4.5/5 (27)