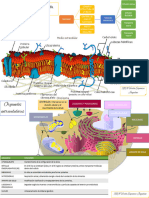
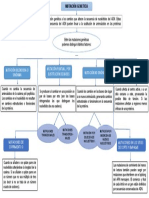
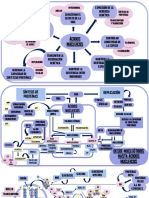
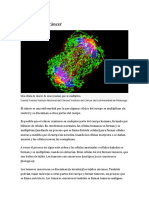
También podría gustarte
- Mapa Conceptual NucleoDocumento2 páginasMapa Conceptual NucleoMiller Noah MichaleAún no hay calificaciones
- Biologia AdnDocumento2 páginasBiologia AdnJasmin ValdiviaAún no hay calificaciones
- Organelos CelularesDocumento19 páginasOrganelos CelularesCristinaAún no hay calificaciones
- Accede A Apuntes, Guías, Libros y Más de Tu Carrera: La Célula Eucariota 1 PagDocumento2 páginasAccede A Apuntes, Guías, Libros y Más de Tu Carrera: La Célula Eucariota 1 PagPaola SilvaAún no hay calificaciones
- Resonancia Magnética Nuclear (RMN) .Documento2 páginasResonancia Magnética Nuclear (RMN) .Rodríguez Torres Jesús OmarAún no hay calificaciones
- BC Cuaderno de Ejercicios 3Documento16 páginasBC Cuaderno de Ejercicios 3camiAún no hay calificaciones
- OrganelosDocumento2 páginasOrganelosfarmaciasanbernardo3Aún no hay calificaciones
- Nucleo InterfasicoDocumento5 páginasNucleo InterfasicoBenjamin TugenderAún no hay calificaciones
- NucleoDocumento1 páginaNucleoGisel elena García mayoralAún no hay calificaciones
- Upao Los Caminates Del NorteDocumento50 páginasUpao Los Caminates Del NorteAlejandra PozoAún no hay calificaciones
- Citologia III Nucleo y CromosomasDocumento32 páginasCitologia III Nucleo y CromosomasNoa NatividadAún no hay calificaciones
- Cuaderno - de - Ejercicios - N°3-BM - 2023 BIOLOGIADocumento32 páginasCuaderno - de - Ejercicios - N°3-BM - 2023 BIOLOGIAValentina Gómez100% (1)
- 943-Cuaderno de Ejercicios N°2 BC 2024Documento17 páginas943-Cuaderno de Ejercicios N°2 BC 2024Maria-jose analia Novoa cidAún no hay calificaciones
- NEUROPLASTICIDADDocumento1 páginaNEUROPLASTICIDADOriana ReyesAún no hay calificaciones
- Sistema de Conduccion Electria Del CorazonDocumento1 páginaSistema de Conduccion Electria Del Corazonapi-472508482Aún no hay calificaciones
- Organizador MembranaDocumento13 páginasOrganizador MembranaLuis Fernando Llacta CiprianoAún no hay calificaciones
- Unidad 7. Transcripcion y Regulacion GenicaDocumento39 páginasUnidad 7. Transcripcion y Regulacion GenicaPablo Ignacio BarriasAún no hay calificaciones
- Actividadessobreestructuracelular1 120318122717 Phpapp02Documento4 páginasActividadessobreestructuracelular1 120318122717 Phpapp02WilliamAndresAlvarezAún no hay calificaciones
- Mapa DiomarDocumento1 páginaMapa DiomarDiana Katherine Vergel PalenciaAún no hay calificaciones
- Como Ventanas Hacia Una Antigua OrganelaDocumento3 páginasComo Ventanas Hacia Una Antigua OrganelaNatalia CasanaAún no hay calificaciones
- NEOPLASIASDocumento3 páginasNEOPLASIASmarAún no hay calificaciones
- ActiDocumento5 páginasActigrupotextilrysAún no hay calificaciones
- Actividad 03 U1 Cuadro ComparativoDocumento3 páginasActividad 03 U1 Cuadro ComparativoKevin RiveraAún no hay calificaciones
- Wa0009.Documento2 páginasWa0009.Jerusalen m.mAún no hay calificaciones
- Infografia Apoptosis y NecrosisDocumento3 páginasInfografia Apoptosis y NecrosisKevin GonzalezAún no hay calificaciones
- DMREDocumento4 páginasDMREalyson089Aún no hay calificaciones
- ActividadessobremembranaDocumento4 páginasActividadessobremembranaCecilia Delgado GaleanoAún no hay calificaciones
- Mapa Conceptual Nucleo CelularDocumento2 páginasMapa Conceptual Nucleo CelularMasciel Herrera VargasAún no hay calificaciones
- InfografiaDocumento1 páginaInfografiaJESICA VIDAL ESCOBARAún no hay calificaciones
- Mapa ConceptualDocumento2 páginasMapa ConceptualEmanuel CarrilloAún no hay calificaciones
- 07-Los Ácidos Nucleicos - Mapa SemánticoDocumento1 página07-Los Ácidos Nucleicos - Mapa SemánticoModesto Chavez100% (1)
- Tabla Musc UlosDocumento2 páginasTabla Musc UlosLuis Antonio Ramírez RosalesAún no hay calificaciones
- Mapa Conceptual Nucleo CelularDocumento2 páginasMapa Conceptual Nucleo CelularAndy Miguel Garcia ChoqueAún no hay calificaciones
- Trastornos de La CoagulacionDocumento56 páginasTrastornos de La CoagulacionJhon Esteban Camacho NocobeAún no hay calificaciones
- Resonancia MagnéticaDocumento1 páginaResonancia MagnéticaRodríguez Torres Jesús OmarAún no hay calificaciones
- Celulas Del Sistema NerviosoDocumento1 páginaCelulas Del Sistema NerviosoclaraAún no hay calificaciones
- Capitulo 4 DXDocumento25 páginasCapitulo 4 DXAngeles JacoboAún no hay calificaciones
- Rimapa Guerrero Clendy - Citogenetica - S01Documento5 páginasRimapa Guerrero Clendy - Citogenetica - S01Clendy Yudith Rimapa GuerreroAún no hay calificaciones
- Aparato de Golgi MapaDocumento3 páginasAparato de Golgi MapaFernando Jose Cahuana MamaniAún no hay calificaciones
- Cuadro ComparativoDocumento5 páginasCuadro ComparativoJEMNI MIRIAM MEZONES CORTÉZAún no hay calificaciones
- Célula ResumenDocumento1 páginaCélula ResumenxochicaleAún no hay calificaciones
- 09 - LacélulaIV 2Documento49 páginas09 - LacélulaIV 2Daniel ZapicoAún no hay calificaciones
- Desarollo Embriológico Del SNDocumento1 páginaDesarollo Embriológico Del SNFabi Vizcarra NorattoAún no hay calificaciones
- Clase Teorica Nucleo CelularDocumento46 páginasClase Teorica Nucleo CelularCristian qc11Aún no hay calificaciones
- Infografia Evolucion Humana Ilustrado Colores NeutralesDocumento2 páginasInfografia Evolucion Humana Ilustrado Colores NeutralesGabriel GonzálezAún no hay calificaciones
- GENESER Histologia 4a Ed 5Documento1 páginaGENESER Histologia 4a Ed 5shine lighAún no hay calificaciones
- Mapa Mental Fases y Puntos de Control Del Ciclo CelularDocumento2 páginasMapa Mental Fases y Puntos de Control Del Ciclo CelularSakura Furinyi100% (1)
- Biologia 3Documento32 páginasBiologia 3Santos Canahuire CondoriAún no hay calificaciones
- CitologíaDocumento14 páginasCitologíaWhili VasAún no hay calificaciones
- Histo PresentaciónDocumento42 páginasHisto PresentaciónMAYKOL Pantoja NicolasAún no hay calificaciones
- MEMBRANA CELULAR Clase 7Documento1 páginaMEMBRANA CELULAR Clase 7NOE CORNE CAMARAAún no hay calificaciones
- Expo Fisiología BDocumento24 páginasExpo Fisiología BBelen Garcia dionicioAún no hay calificaciones
- Infografía HistologíaDocumento2 páginasInfografía Histologíalaura2005.acAún no hay calificaciones
- Morfo Embrio Semana 2 - Resumen UpaoDocumento4 páginasMorfo Embrio Semana 2 - Resumen UpaoTomas CabanillasAún no hay calificaciones
- Biologia Clase Ammed 2Documento21 páginasBiologia Clase Ammed 2Escarlet MorenoAún no hay calificaciones
- Tablero en BlancoDocumento1 páginaTablero en Blancokatherinejojoa0Aún no hay calificaciones
- Renovación, Migración y Diferenciación de Las CélulasDocumento15 páginasRenovación, Migración y Diferenciación de Las CélulasAlejandro Chung BenitesAún no hay calificaciones
- TPE3 - Introducción-BiologíaDocumento15 páginasTPE3 - Introducción-BiologíasofiAún no hay calificaciones
- Ácidos Nucleicos y Nucleótidos. FuncionesDocumento5 páginasÁcidos Nucleicos y Nucleótidos. FuncionesArturo VelazcoAún no hay calificaciones
- Introducción a la Biología: RESÚMENES UNIVERSITARIOSDe EverandIntroducción a la Biología: RESÚMENES UNIVERSITARIOSCalificación: 5 de 5 estrellas5/5 (1)
- 7-Dna VDocumento5 páginas7-Dna VCamila Gonzalez PiatAún no hay calificaciones
- Ley de CoulombDocumento8 páginasLey de CoulombCamila Gonzalez PiatAún no hay calificaciones
- 5-Dna IiiDocumento9 páginas5-Dna IiiCamila Gonzalez PiatAún no hay calificaciones
- Acuerdo 3354Documento40 páginasAcuerdo 3354Camila Gonzalez PiatAún no hay calificaciones
- Concursos y Quiebras 12Documento25 páginasConcursos y Quiebras 12Camila Gonzalez PiatAún no hay calificaciones
- Resumen Practico PDFDocumento99 páginasResumen Practico PDFCamila Gonzalez PiatAún no hay calificaciones
- 8.técnicas Básicas en Biología MolecularDocumento41 páginas8.técnicas Básicas en Biología MolecularVianney CervantesAún no hay calificaciones
- Guia 2Documento6 páginasGuia 2Marylin SanchezAún no hay calificaciones
- 1era Clase de Biología CelularDocumento76 páginas1era Clase de Biología CelularMaria Belen MartinezAún no hay calificaciones
- Tarea de Practica Sobre NucleotidosDocumento4 páginasTarea de Practica Sobre NucleotidosPrime VideoAún no hay calificaciones
- Barbilla PartidaDocumento2 páginasBarbilla PartidahugoAún no hay calificaciones
- Acidos Nucleicos 17-18-170910104832 PDFDocumento17 páginasAcidos Nucleicos 17-18-170910104832 PDFAnonymous GS7buYAún no hay calificaciones
- Marchitez de La PiñaDocumento14 páginasMarchitez de La PiñaBetssy Amparo Basurto RomeroAún no hay calificaciones
- Practico Nuevo 12 Hardy W 2017Documento8 páginasPractico Nuevo 12 Hardy W 2017DarioCastroAún no hay calificaciones
- 1.-Unidad I Sesión I Enunciados Problemas ClaseDocumento2 páginas1.-Unidad I Sesión I Enunciados Problemas Clase01001101Aún no hay calificaciones
- 1.c.n.noveno Primer Periodo-Guia UnoDocumento22 páginas1.c.n.noveno Primer Periodo-Guia UnoIsa JaramilloAún no hay calificaciones
- Guía Ciclo CelularDocumento1 páginaGuía Ciclo CelularElianetteAún no hay calificaciones
- Problemas Sobre Herencia Ligada Al SexoDocumento9 páginasProblemas Sobre Herencia Ligada Al SexoJonathan RamosAún no hay calificaciones
- García Karo, Tamara Flor (2014) - Manipulación Genética en Humanos Transformación o Mutación Potencial Riesgo para La Diversidad de (... )Documento6 páginasGarcía Karo, Tamara Flor (2014) - Manipulación Genética en Humanos Transformación o Mutación Potencial Riesgo para La Diversidad de (... )Isaac Fernandez RoblesAún no hay calificaciones
- Mutaciones en Word para ImprimirDocumento7 páginasMutaciones en Word para ImprimirPercy Neyra100% (1)
- GAMETOGÉNESISDocumento21 páginasGAMETOGÉNESISAngelah Katherine Q ChAún no hay calificaciones
- Estructura y Funcion de Acidos Nucleicos IDocumento34 páginasEstructura y Funcion de Acidos Nucleicos IChristian MartinoniAún no hay calificaciones
- Soluciones Examen de Biología y Geología de 4º ESO 21-3-2012Documento5 páginasSoluciones Examen de Biología y Geología de 4º ESO 21-3-2012jccambronero63% (8)
- Bioquímica - Parcial 1 - Preguntas de Duplicación y TranscripciónDocumento3 páginasBioquímica - Parcial 1 - Preguntas de Duplicación y TranscripciónTahía TamaraAún no hay calificaciones
- Practica # 1 Las MutacionesDocumento8 páginasPractica # 1 Las MutacionesRania ArbajeAún no hay calificaciones
- Extraccion de Adn A Partir de Sangre Periferica 1Documento4 páginasExtraccion de Adn A Partir de Sangre Periferica 1PamelaaGaytan100% (1)
- Principios de La GeneticaDocumento67 páginasPrincipios de La GeneticaAmiel Mendoza HuamaniAún no hay calificaciones
- Sindrome de EdwardsDocumento9 páginasSindrome de EdwardsArtis ValyeAún no hay calificaciones
- Línea Del Tiempo Biología MolecularDocumento1 páginaLínea Del Tiempo Biología MolecularAnell Morales Samuel100% (2)
- ¿Qué Es El ADN MedlinePlus GeneticsDocumento1 página¿Qué Es El ADN MedlinePlus GeneticsThiare AntoniaAún no hay calificaciones
- Ticket de Salida Ciclo CelularDocumento2 páginasTicket de Salida Ciclo CelularYoselyn CornejoAún no hay calificaciones
- Quien Fue Gregor MendelDocumento5 páginasQuien Fue Gregor Mendelfranni carapaicaAún no hay calificaciones
- AcondroplasiaDocumento6 páginasAcondroplasiaRosmary OrtizAún no hay calificaciones
- Extraccion de Adn en Celulas VegetalesDocumento16 páginasExtraccion de Adn en Celulas VegetalesLizHernandez100% (1)
- Definición Del CáncerDocumento28 páginasDefinición Del CáncerGuisel MenchúAún no hay calificaciones
- Genetica 1.2 ParcialDocumento13 páginasGenetica 1.2 ParcialPaulo CarvalhoAún no hay calificaciones