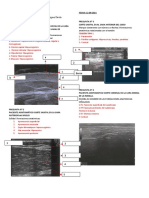
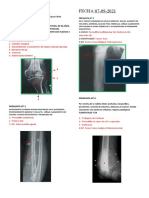
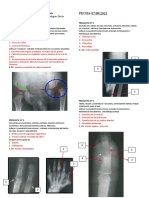
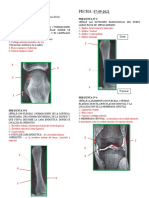
También podría gustarte
- Guía Practica #15Documento1 páginaGuía Practica #15Cynthia Miranda SaldañaAún no hay calificaciones
- Guía Practica #13Documento1 páginaGuía Practica #13Cynthia Miranda Saldaña100% (1)
- Guía Practica #14Documento1 páginaGuía Practica #14Cynthia Miranda SaldañaAún no hay calificaciones
- Guía Practica #20Documento1 páginaGuía Practica #20Cynthia Miranda SaldañaAún no hay calificaciones
- Guía Practica #11Documento2 páginasGuía Practica #11Cynthia Miranda Saldaña0% (1)
- Guía Practica #17Documento1 páginaGuía Practica #17Cynthia Miranda SaldañaAún no hay calificaciones
- Guía Practica #18Documento1 páginaGuía Practica #18Cynthia Miranda SaldañaAún no hay calificaciones
- Guia Practica #1Documento1 páginaGuia Practica #1Cynthia Miranda SaldañaAún no hay calificaciones
- Guia Practica #2Documento1 páginaGuia Practica #2Cynthia Miranda SaldañaAún no hay calificaciones
- Guia Practica #3Documento1 páginaGuia Practica #3Cynthia Miranda Saldaña100% (1)
- Guia Practica #8Documento1 páginaGuia Practica #8Cynthia Miranda SaldañaAún no hay calificaciones
- Guia Practica #9Documento1 páginaGuia Practica #9Cynthia Miranda SaldañaAún no hay calificaciones
- Anemias MegaloblásticasDocumento10 páginasAnemias MegaloblásticasCynthia Miranda SaldañaAún no hay calificaciones
- La Interacción Entre Las Plaquetas y La CoagulaciónDocumento8 páginasLa Interacción Entre Las Plaquetas y La CoagulaciónCynthia Miranda SaldañaAún no hay calificaciones
- Guía Practica #7Documento1 páginaGuía Practica #7Cynthia Miranda SaldañaAún no hay calificaciones
- Guia Practica #6Documento1 páginaGuia Practica #6Cynthia Miranda SaldañaAún no hay calificaciones
- Diagnostico Limitrofe Neurologia PsiquiatriaDocumento18 páginasDiagnostico Limitrofe Neurologia PsiquiatriaHumberto Castillo MartellAún no hay calificaciones
- Folleto Biomateriales Todos Campaña New 490.561-Es - LowDocumento80 páginasFolleto Biomateriales Todos Campaña New 490.561-Es - LowGolden VirginiaAún no hay calificaciones
- Reparacion TisularDocumento7 páginasReparacion Tisularalma monserrat bass vargasAún no hay calificaciones
- Células MadreDocumento46 páginasCélulas MadreMartín Vuelta100% (1)
- SabiiaDocumento807 páginasSabiiaAnonymous 0lxTug67Aún no hay calificaciones
- Regeneracion Tisular GuiadaDocumento1 páginaRegeneracion Tisular GuiadaDaynor Churqui VargasAún no hay calificaciones
- Práctica 9 de PatologíaDocumento7 páginasPráctica 9 de Patologíafrida ornelasAún no hay calificaciones
- Cirugía Regenerativa Perirradicular en Un Incisivo Central Superior - En.esDocumento7 páginasCirugía Regenerativa Perirradicular en Un Incisivo Central Superior - En.esAngela RamirezAún no hay calificaciones
- CaratulaDocumento8 páginasCaratulafernando haroAún no hay calificaciones
- Informe de Axolotl... Mi SalamandraDocumento13 páginasInforme de Axolotl... Mi SalamandraNoelia Rousdary Burgoa ApazaAún no hay calificaciones
- Pez CebraDocumento6 páginasPez CebraGabrielCamarena100% (1)
- Jhon AlvarezDocumento15 páginasJhon AlvarezJ ArmorAún no hay calificaciones
- Reparación de Tejido Blando: Resultado de Aprendizaje / ContenidosDocumento22 páginasReparación de Tejido Blando: Resultado de Aprendizaje / ContenidosMaite RodriguezAún no hay calificaciones
- 131-Texto Del Artículo-514-1-10-20150826Documento11 páginas131-Texto Del Artículo-514-1-10-20150826Alejandra Villavicencio FloresAún no hay calificaciones
- Reparación de TejidosDocumento8 páginasReparación de TejidosInésAún no hay calificaciones
- Cicatrices de Acne y Regenaracion de Las CelulasDocumento50 páginasCicatrices de Acne y Regenaracion de Las CelulasMathias Cabarcas FrancoAún no hay calificaciones
- Solemne 2 TOSFA UNAB ViñaDocumento63 páginasSolemne 2 TOSFA UNAB ViñaFernanda M Arroyo OrtizAún no hay calificaciones
- Led 7 ColoresDocumento2 páginasLed 7 ColoresAlina Cecilia SosaAún no hay calificaciones
- Hammarstrom1997 en EsDocumento9 páginasHammarstrom1997 en EsHector nemysan Murillo SotoAún no hay calificaciones
- Suspensiones de Células Embriogénicas de Banano y Plátano 946Documento36 páginasSuspensiones de Células Embriogénicas de Banano y Plátano 946Anonymous KbTL8xAún no hay calificaciones
- Bioestimulacion Cutanea Con Plasma EnriqDocumento24 páginasBioestimulacion Cutanea Con Plasma Enriqpaola poppeAún no hay calificaciones
- Regeneración Natural Del Bosque Seco de La Provincia de Loja PDFDocumento13 páginasRegeneración Natural Del Bosque Seco de La Provincia de Loja PDFZhofre Aguirre MendozaAún no hay calificaciones
- Atlas de Pato PUCDocumento432 páginasAtlas de Pato PUCuzume278Aún no hay calificaciones
- Software de Control Del Tren Motriz F-550Documento6 páginasSoftware de Control Del Tren Motriz F-550Carlos LagoAún no hay calificaciones
- Regeneration Hepatic - En.esDocumento13 páginasRegeneration Hepatic - En.esMaría De los SantosAún no hay calificaciones
- Monografia Viagland Espa OlDocumento63 páginasMonografia Viagland Espa Oljotasantos6Aún no hay calificaciones
- Factores de Crecimiento y Regeneración RenalDocumento2 páginasFactores de Crecimiento y Regeneración Renalcielo santiago barrios angelesAún no hay calificaciones
- Yañez-Espinoza, 2017. Adaptación de Las Plantas LeñosasDocumento52 páginasYañez-Espinoza, 2017. Adaptación de Las Plantas LeñosasDaniel DUperonAún no hay calificaciones
- CuracionesDocumento45 páginasCuracionesAnonymous AbGW1XQOeAún no hay calificaciones
- MFPH 1 Semana 7Documento3 páginasMFPH 1 Semana 7Henry DuqueAún no hay calificaciones