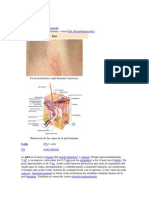
También podría gustarte
- Monografia de Aves de MéxicoDocumento43 páginasMonografia de Aves de MéxicoMario Lara100% (1)
- Histología y Embrología del Sitema NerviosoDe EverandHistología y Embrología del Sitema NerviosoCalificación: 4 de 5 estrellas4/5 (4)
- Cuadernillo Aves ArgentinasDocumento32 páginasCuadernillo Aves Argentinasjoe1974100% (2)
- Definición de AviculturaDocumento5 páginasDefinición de AviculturaMariluzLeiva100% (1)
- Arte de Acampar I - CampamentismoDocumento17 páginasArte de Acampar I - CampamentismoJ Elvis SalazarAún no hay calificaciones
- Desarrollo Ontogenico Del Sistema NerviosoDocumento18 páginasDesarrollo Ontogenico Del Sistema NerviosoLorena100% (1)
- Las Adaptaciones Al AmbienteDocumento31 páginasLas Adaptaciones Al Ambientelourdes100% (1)
- Rehabilitacion de Aves Salvajes HeridasDocumento181 páginasRehabilitacion de Aves Salvajes HeridasSoyKlicheRodriguezAún no hay calificaciones
- FenologíaDocumento213 páginasFenologíaJhair Alexander Elías LamadridAún no hay calificaciones
- La GlobalizaciónDocumento19 páginasLa GlobalizaciónJhair Alexander Elías LamadridAún no hay calificaciones
- Kolisko, Eugen - Zoología para TodosDocumento45 páginasKolisko, Eugen - Zoología para Todossarai Martinez100% (2)
- Savage Island 01 El Corazón de Savage BookDocumento156 páginasSavage Island 01 El Corazón de Savage BookMcChesney_PasseyAún no hay calificaciones
- Sistema TegumentarioDocumento12 páginasSistema TegumentarioandresJB12Aún no hay calificaciones
- Resumen de El Proceso de Hominización: RESÚMENES UNIVERSITARIOSDe EverandResumen de El Proceso de Hominización: RESÚMENES UNIVERSITARIOSAún no hay calificaciones
- Tema 11-Bloque II-La Piel. Estructura y FuncionesDocumento7 páginasTema 11-Bloque II-La Piel. Estructura y FuncionesGenesis Gonzalez ReyesAún no hay calificaciones
- Narracion Oral de CuentosDocumento3 páginasNarracion Oral de Cuentosrosana100% (1)
- Que Es La PielDocumento8 páginasQue Es La PielBiomagnetista Experta EN Masaje Reductivos100% (1)
- Animales Vertebrados e InvertebradosDocumento23 páginasAnimales Vertebrados e InvertebradosErika Calderon Riquelme0% (1)
- S6 Piel y AnexosDocumento6 páginasS6 Piel y AnexosDiego Peña RomeroAún no hay calificaciones
- Antologà - As Anat I 2023-2 OkDocumento55 páginasAntologà - As Anat I 2023-2 OkAnna Esther MollhoffAún no hay calificaciones
- La PielDocumento9 páginasLa PielDorysbell NuñezAún no hay calificaciones
- CARSON Capitulo de Epidermis PDFDocumento13 páginasCARSON Capitulo de Epidermis PDFSofia TorresAún no hay calificaciones
- Desarrollo y Estructura de La PielDocumento14 páginasDesarrollo y Estructura de La PielAlba Noemi MorenoAún no hay calificaciones
- Mutaciones EspontáneasDocumento6 páginasMutaciones EspontáneasJilber Ramon Sanchez VicentAún no hay calificaciones
- TSCSalud2 Unidad3Documento30 páginasTSCSalud2 Unidad3naranjito2222Aún no hay calificaciones
- Embriologia Alteraciones en El Sistema TegumentarioDocumento20 páginasEmbriologia Alteraciones en El Sistema TegumentarioDavid VillaAún no hay calificaciones
- Endogenesis PDFDocumento6 páginasEndogenesis PDFFranyeli SarmientoAún no hay calificaciones
- Sitema TegumentoDocumento11 páginasSitema TegumentoJozé Christhian Cornejo PérezAún no hay calificaciones
- OrganogenesisDocumento5 páginasOrganogenesiskatybaluz777Aún no hay calificaciones
- Capas Blastodérmicas ENSAYODocumento4 páginasCapas Blastodérmicas ENSAYOLeonel Ciprian ArrietaAún no hay calificaciones
- El TegumentoDocumento38 páginasEl Tegumentoeduardojr1234Aún no hay calificaciones
- FECUNDACIONDocumento2 páginasFECUNDACIONJosé ContrerasAún no hay calificaciones
- Piel CorderoDocumento86 páginasPiel CorderoAndreaCostales100% (1)
- Caso Clinico PielDocumento4 páginasCaso Clinico PielnathalyAún no hay calificaciones
- Carlson Sistema TegumentarioDocumento10 páginasCarlson Sistema TegumentarioMiguel Angel LopezAún no hay calificaciones
- Tarea Sesión de Aprendizaje N9Documento11 páginasTarea Sesión de Aprendizaje N9Henry Yamil Martinez MartinezAún no hay calificaciones
- Embriología Del Sistema TegumentarioDocumento12 páginasEmbriología Del Sistema TegumentarioLG Zuri100% (1)
- Anatomia y Fisiologia de La PielDocumento13 páginasAnatomia y Fisiologia de La PielPierina Anthonella Farias SuarezAún no hay calificaciones
- Derivados Mesodermales Histogénesis Del Tejido Conectivo y Muscular 11Documento4 páginasDerivados Mesodermales Histogénesis Del Tejido Conectivo y Muscular 11ddiazc231Aún no hay calificaciones
- 1 Embriologia Del Sistema TegumentarioDocumento11 páginas1 Embriologia Del Sistema TegumentarioNoeRZAEAún no hay calificaciones
- Tegu HistologiaDocumento66 páginasTegu HistologiaEduardo TovarAún no hay calificaciones
- CUESTIONARIO N5 de EnbriologiaDocumento4 páginasCUESTIONARIO N5 de EnbriologiaAnyi LopezAún no hay calificaciones
- Desarrollo EmbrionarioDocumento5 páginasDesarrollo EmbrionarioCALVO LIMONCHE SILVIA, TURNO VESPERTINOAún no hay calificaciones
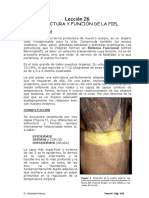
- Lección 26 TEXTO1-PielDocumento16 páginasLección 26 TEXTO1-PielDavid Del Pozo MerinoAún no hay calificaciones
- Unidad 3Documento25 páginasUnidad 3AndresReyesDominguez0% (1)
- Glosario de Términos - Angeline Hurel Laina 2-5Documento12 páginasGlosario de Términos - Angeline Hurel Laina 2-5Angeline HurelAún no hay calificaciones
- Piel - Wikipedia, La Enciclopedia LibreDocumento74 páginasPiel - Wikipedia, La Enciclopedia Librenataly cepedaAún no hay calificaciones
- Capitulo 7 Tegumento PDFDocumento24 páginasCapitulo 7 Tegumento PDFMarlon Ortiz Hummer100% (2)
- TEJIDO EPITELIAL y TEJIDO TEGUMENTARIODocumento6 páginasTEJIDO EPITELIAL y TEJIDO TEGUMENTARIOKaren UsecheAún no hay calificaciones
- Desarrollo Embrionario - Etapas y Sus Características (Semana A Semana) - LifederDocumento15 páginasDesarrollo Embrionario - Etapas y Sus Características (Semana A Semana) - LifederMevis Paola Ruiz JuncoAún no hay calificaciones
- Ensayo Sobre La Piel (Semiologia)Documento16 páginasEnsayo Sobre La Piel (Semiologia)Oc gamesAún no hay calificaciones
- Capa GerminalDocumento5 páginasCapa GerminalWrobercito CutipaAún no hay calificaciones
- Gastrulacion RaipDocumento6 páginasGastrulacion RaipRicardo Iñiguez ParraAún no hay calificaciones
- ESPOCHDocumento33 páginasESPOCHSantiago MartinezAún no hay calificaciones
- Notas Sobre FecundacionDocumento6 páginasNotas Sobre FecundacionFranyer VargasAún no hay calificaciones
- Ensayo Sobre La Piel (Semiologia) - 1Documento16 páginasEnsayo Sobre La Piel (Semiologia) - 1Oc gamesAún no hay calificaciones
- Aparatos y Sistemas Derivados de Las Tres Hojas EmbrionariasDocumento5 páginasAparatos y Sistemas Derivados de Las Tres Hojas Embrionariasfrigo sumi cajaAún no hay calificaciones
- Biologia Del Desarrollo 1Documento76 páginasBiologia Del Desarrollo 1BIOLOGIA UNIPAMPLONAAún no hay calificaciones
- 4 Dif LGR Diferenciacion CelularDocumento2 páginas4 Dif LGR Diferenciacion CelularHumberto Bustos CalabaceroAún no hay calificaciones
- Histología de La Piel FinalDocumento25 páginasHistología de La Piel Finalkarina corporanAún no hay calificaciones
- El EctodermoDocumento4 páginasEl EctodermoNATALIA ALEMANAún no hay calificaciones
- BIOLOGIA Silvana GuilcapiDocumento7 páginasBIOLOGIA Silvana GuilcapiDaniel Rosero100% (1)
- TAREA 2 Neuroanatomia FuncionalDocumento4 páginasTAREA 2 Neuroanatomia Funcionalhinojosalevi22Aún no hay calificaciones
- Desarrollo EmbrionarioDocumento3 páginasDesarrollo EmbrionarioThiago VelazquezAún no hay calificaciones
- Epitelio Apunte 10Documento38 páginasEpitelio Apunte 10Gustavo AlbertoAún no hay calificaciones
- GJGNDocumento4 páginasGJGNROSA FAILOCAún no hay calificaciones
- Capitulo I Anatomia y Fisiologia de La PielDocumento51 páginasCapitulo I Anatomia y Fisiologia de La PielAarón E. CaríasAún no hay calificaciones
- OrganogénesisDocumento5 páginasOrganogénesisjovmac2uAún no hay calificaciones
- Caso DermatofuncionalDocumento13 páginasCaso DermatofuncionalAlejandra RomeroAún no hay calificaciones
- Inmunología de la piel: Segunda ediciónDe EverandInmunología de la piel: Segunda ediciónAún no hay calificaciones
- Recurso Alternativo Lluvia Acida Causas y ConsecuenciasDocumento4 páginasRecurso Alternativo Lluvia Acida Causas y ConsecuenciasJhair Alexander Elías LamadridAún no hay calificaciones
- Coportamiento AgonísticoDocumento22 páginasCoportamiento AgonísticoJhair Alexander Elías LamadridAún no hay calificaciones
- Informe Bienestar AnimalDocumento3 páginasInforme Bienestar AnimalJhair Alexander Elías Lamadrid0% (1)
- Trematodes y CestodesDocumento30 páginasTrematodes y CestodesJhair Alexander Elías LamadridAún no hay calificaciones
- Desarrollo de Una PapayaDocumento16 páginasDesarrollo de Una PapayaJhair Alexander Elías LamadridAún no hay calificaciones
- 198132376Documento87 páginas198132376JEFFERSON JHAIRO ORE GENEBROSOAún no hay calificaciones
- La CreacionDocumento635 páginasLa Creacionariahnna2013100% (1)
- Diamante Gould INODocumento4 páginasDiamante Gould INOCANARIL NN100% (1)
- Manual de Identificacic3b3n en El Campo Del C3a1guila Real Mediterrc3a1nea 1Documento13 páginasManual de Identificacic3b3n en El Campo Del C3a1guila Real Mediterrc3a1nea 1kkupus.14Aún no hay calificaciones
- El Casco de Los EquinosDocumento8 páginasEl Casco de Los Equinosmacarenah1994Aún no hay calificaciones
- Gallos y OtrosDocumento139 páginasGallos y OtrosJairo GonzalezAún no hay calificaciones
- M'Os Aerobios Mesofilos ViablesDocumento27 páginasM'Os Aerobios Mesofilos ViablesErick CQAún no hay calificaciones
- Rejuvenecer Como El ÁguilaDocumento5 páginasRejuvenecer Como El ÁguilaJoe Pad50% (2)
- Evaluación de Lenguaje Segunda UnidadDocumento2 páginasEvaluación de Lenguaje Segunda UnidadMary Herrera0% (1)
- 2°. El CasuarioDocumento2 páginas2°. El CasuarioAnahí TropiniAún no hay calificaciones
- Cedulas Aves Bastidor 2Documento18 páginasCedulas Aves Bastidor 2Roberto Carlos Calderón CarvajalAún no hay calificaciones
- Harina de SangreDocumento4 páginasHarina de SangreNarcisa LoorAún no hay calificaciones
- Actividad 4 Continuidad Pedagogica (Abril-Mayo)Documento20 páginasActividad 4 Continuidad Pedagogica (Abril-Mayo)oliverroxana4Aún no hay calificaciones
- 1° - Ciencias - 8TIPOS CUBIERTASDocumento5 páginas1° - Ciencias - 8TIPOS CUBIERTASSussanna GomezAún no hay calificaciones
- Tarea N°1Documento3 páginasTarea N°1Marianela JaureAún no hay calificaciones
- Clase 2 Razas y LÃ-neasDocumento89 páginasClase 2 Razas y LÃ-neasCV YulisaAún no hay calificaciones
- AvesDocumento4 páginasAvesmossoulAún no hay calificaciones
- Anatomia Externa de Las AvesDocumento7 páginasAnatomia Externa de Las Avesdavid quinterosAún no hay calificaciones
- Avicola Unido PDFDocumento86 páginasAvicola Unido PDFIgnacio MonsalvoAún no hay calificaciones
- 4o Grado. Cuadernillo Aprende en Casa. Abril 2021Documento38 páginas4o Grado. Cuadernillo Aprende en Casa. Abril 2021JuanBondAún no hay calificaciones