También podría gustarte
- Aparato Respiratorio GallinaDocumento8 páginasAparato Respiratorio GallinayvanavalerogomezAún no hay calificaciones
- 10 ELEC BL Estrés y Drogas - 2015Documento12 páginas10 ELEC BL Estrés y Drogas - 2015Maka Gomez Ibarra0% (1)
- Cuestionario PoscosechaDocumento8 páginasCuestionario PoscosechaKaren MoraAún no hay calificaciones
- DrogasDocumento6 páginasDrogasMaka Gomez IbarraAún no hay calificaciones
- Hormonas y SexualidadDocumento12 páginasHormonas y SexualidadMaka Gomez IbarraAún no hay calificaciones
- BM25 Ecología IIIDocumento20 páginasBM25 Ecología IIIMaka Gomez IbarraAún no hay calificaciones
- Sinapsis y NeurotransmisoresDocumento2 páginasSinapsis y NeurotransmisoresMaka Gomez IbarraAún no hay calificaciones
- Niveles de Organizacion PDFDocumento1 páginaNiveles de Organizacion PDFMaka Gomez IbarraAún no hay calificaciones
- Redes Alimentarias PDFDocumento1 páginaRedes Alimentarias PDFMaka Gomez IbarraAún no hay calificaciones
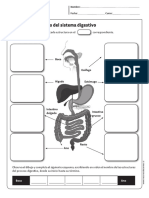
- Sistema Digestivo PDFDocumento1 páginaSistema Digestivo PDFMaka Gomez IbarraAún no hay calificaciones
- Guía de Sistema Reproductor MasculinoDocumento2 páginasGuía de Sistema Reproductor MasculinoMaka Gomez Ibarra100% (1)
- Selección Múltiple - CélulaDocumento2 páginasSelección Múltiple - CélulaKro Fuentes Leiva100% (2)
- Prueba Propiedades y Leyes de Los GasesDocumento3 páginasPrueba Propiedades y Leyes de Los GasesMaka Gomez Ibarra100% (1)
- Guia Redox 4ºA MEDIO BASICODocumento4 páginasGuia Redox 4ºA MEDIO BASICOMaka Gomez IbarraAún no hay calificaciones
- Clase 7 Fisiología Muscular y Ventilación 2015Documento44 páginasClase 7 Fisiología Muscular y Ventilación 2015Maka Gomez Ibarra0% (1)
- Clase 6 Receptores El Globo Ocular 2015Documento32 páginasClase 6 Receptores El Globo Ocular 2015Maka Gomez Ibarra0% (1)
- Clase de TiroidesDocumento68 páginasClase de TiroidesyhismeldAún no hay calificaciones
- Ejercicios Q BartonDocumento7 páginasEjercicios Q BartonTatto RamirezAún no hay calificaciones
- Histologia Silabo 2017-IIDocumento11 páginasHistologia Silabo 2017-IIARis Huayllani GavilanAún no hay calificaciones
- Nen. BiomagnetismoDocumento33 páginasNen. BiomagnetismoTxell Martínez AmperiAún no hay calificaciones
- Generalidades y Fundamentos Del AguaDocumento12 páginasGeneralidades y Fundamentos Del AguaJulio David Guzmán BenítezAún no hay calificaciones
- She Becomes Glamorous After The Engagement Annulment 401-465Documento242 páginasShe Becomes Glamorous After The Engagement Annulment 401-465Marian CAún no hay calificaciones
- APIDocumento6 páginasAPIroyer944Aún no hay calificaciones
- Tecnologias Emergentes en AlimentosDocumento40 páginasTecnologias Emergentes en AlimentosKatheryn PeraltaAún no hay calificaciones
- Empaque NanotecnologiaDocumento25 páginasEmpaque NanotecnologiaAngela RiveroAún no hay calificaciones
- Epicocum, Cladosporium, Mucor y Saprolegnia PDFDocumento29 páginasEpicocum, Cladosporium, Mucor y Saprolegnia PDFHelcib De Los AngelesAún no hay calificaciones
- Biomimetica PDFDocumento8 páginasBiomimetica PDFDaniel UlloaAún no hay calificaciones
- Planteamientos SocialesDocumento1 páginaPlanteamientos SocialesJoaquin IAún no hay calificaciones
- FLAVONOIDESDocumento11 páginasFLAVONOIDESJose AvalosAún no hay calificaciones
- Sesion 3 - Ejercicios Activos y Pasivos, Hidroterapia, TermoterapiaDocumento34 páginasSesion 3 - Ejercicios Activos y Pasivos, Hidroterapia, TermoterapiaJeanet FelixAún no hay calificaciones
- Diario de Campo AdultezDocumento7 páginasDiario de Campo AdultezErika Lizeth PAREDES CASTILLOAún no hay calificaciones
- BilateradosDocumento7 páginasBilateradosDredeltoAún no hay calificaciones
- Edad y Crecimiento BP 2019Documento33 páginasEdad y Crecimiento BP 2019Vladimir SorianoAún no hay calificaciones
- MFM Medicion Funcion Motora Enf Neuromusculares LIBRO Es 20071121114642Documento56 páginasMFM Medicion Funcion Motora Enf Neuromusculares LIBRO Es 20071121114642Patricia Carolina Acosta AgredaAún no hay calificaciones
- Investigacion de Psicologia Social y Comunitaria-1Documento18 páginasInvestigacion de Psicologia Social y Comunitaria-1Blanca CalderónAún no hay calificaciones
- Caracterizacion-De Tamaño de ParticulasDocumento51 páginasCaracterizacion-De Tamaño de ParticulasLizana AdielAún no hay calificaciones
- Plan de Manejo PRM Volcan San PedroDocumento56 páginasPlan de Manejo PRM Volcan San Pedroapi-3758038100% (3)
- Jerarquía de Necesidades, Desarrollando Habilidades de ComunicaciónDocumento7 páginasJerarquía de Necesidades, Desarrollando Habilidades de ComunicaciónJhons D. RamirezAún no hay calificaciones
- Final Genetica Só DR AtuaisDocumento73 páginasFinal Genetica Só DR AtuaisIzabella TeruyaAún no hay calificaciones
- Introducción A La HistologíaDocumento30 páginasIntroducción A La HistologíaRomiglio AlejandroAún no hay calificaciones
- Principios Generales de Análisis e Investigación en Bioquímica Clínica - BioquímicaDocumento9 páginasPrincipios Generales de Análisis e Investigación en Bioquímica Clínica - BioquímicaMaria Renee Lopez VillarroelAún no hay calificaciones
- De La Transmisión Tradente A La Actualización de PosibilidadesDocumento7 páginasDe La Transmisión Tradente A La Actualización de PosibilidadesChitomonoAún no hay calificaciones
- Toledo y Gonzalez de Molina Metabolismo SocialDocumento23 páginasToledo y Gonzalez de Molina Metabolismo SocialangelaAún no hay calificaciones
- Que Es Ecosistema TerrestreDocumento4 páginasQue Es Ecosistema TerrestreGilmer TalaAún no hay calificaciones