También podría gustarte
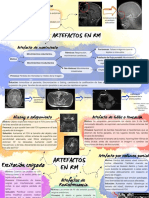
- Esquema Artefactos Resonancia MagnéticaDocumento3 páginasEsquema Artefactos Resonancia MagnéticaAna Cristina Jiménez López100% (1)
- IRM Hecha Facil (... Bueno, Casi)Documento126 páginasIRM Hecha Facil (... Bueno, Casi)MARCELO IVÃ�N FIGUEROA FERREIRAAún no hay calificaciones
- Sopa de Letras SSTDocumento1 páginaSopa de Letras SSTSAÚL MARTÍNEZ VEGA67% (3)
- Clase CardioDocumento58 páginasClase CardioHelen Cid Acevedo100% (1)
- Resonancia MagneticaDocumento16 páginasResonancia MagneticaAntonio VincesAún no hay calificaciones
- Sustancias de Contraste en IRMDocumento33 páginasSustancias de Contraste en IRMAlexisPortugalVillaAún no hay calificaciones
- Fundamentos Físicos de La RMDocumento13 páginasFundamentos Físicos de La RMmiriamAún no hay calificaciones
- Tipos de Secuencias RMDocumento17 páginasTipos de Secuencias RMDoloresAún no hay calificaciones
- Resonancia MagneticaDocumento4 páginasResonancia MagneticaMilagros Condori RamosAún no hay calificaciones
- Secuencias. Spin y Gradiente Eco. Grupo6Documento32 páginasSecuencias. Spin y Gradiente Eco. Grupo6Anahí Ortiz100% (1)
- Urografía ExcretoraDocumento29 páginasUrografía ExcretoraYurian Yaneth Diaz100% (4)
- Resonancia Magnética-3Documento20 páginasResonancia Magnética-3Balumba693Aún no hay calificaciones
- Estudio de Caso - Resonancia MagneticaDocumento8 páginasEstudio de Caso - Resonancia Magneticanatalia holguinAún no hay calificaciones
- Pielografia ExpoDocumento31 páginasPielografia ExpoCyc Ximena0% (1)
- Angioresonancia CarotideaDocumento10 páginasAngioresonancia Carotideajose arayAún no hay calificaciones
- Resonancia MagneticaDocumento136 páginasResonancia Magneticaluislg12100% (3)
- Estudios Contrastados - Digestivo Bajo TIDDocumento83 páginasEstudios Contrastados - Digestivo Bajo TIDCeleste_FeigelAún no hay calificaciones
- Resonancia MagneticaDocumento48 páginasResonancia MagneticaKristian RamirezAún no hay calificaciones
- Resonancia Magnetica en La MedicinaDocumento4 páginasResonancia Magnetica en La MedicinaAndres Esteban GAún no hay calificaciones
- Tecnica RadiograficaDocumento63 páginasTecnica RadiograficaisabellaAún no hay calificaciones
- Magnetizacion TransversalDocumento1 páginaMagnetizacion TransversalNerea Berdun100% (1)
- Resumen UltrasonidosDocumento9 páginasResumen Ultrasonidosdraven8708Aún no hay calificaciones
- CISTOURETROGRAFÍADocumento2 páginasCISTOURETROGRAFÍAyadirandresbairon100% (2)
- Conceptos de RadiologíaDocumento121 páginasConceptos de RadiologíaMarbel CreusAún no hay calificaciones
- Resonancia MagnÉticaDocumento50 páginasResonancia MagnÉticaALE X RAY100% (1)
- ArtefactosDocumento14 páginasArtefactosJeny RamírezAún no hay calificaciones
- AngioresonanciaDocumento5 páginasAngioresonanciacerdocapitalistaAún no hay calificaciones
- Medios de ContrasteDocumento13 páginasMedios de ContrasteNubia GonzalezAún no hay calificaciones
- Capitulo I Fisica Del UltrasonidoDocumento15 páginasCapitulo I Fisica Del UltrasonidoSergio Morales100% (3)
- Estudios Contrastados de Esófago, Estómago y DuodenoDocumento44 páginasEstudios Contrastados de Esófago, Estómago y DuodenoAna Paula Huayta Vizconde50% (2)
- Tema Ecografía-FranDocumento30 páginasTema Ecografía-FranFrancisco Antonio Martinez SerranoAún no hay calificaciones
- CHILERAD Protocolos RM Osteoarticular EESSDocumento11 páginasCHILERAD Protocolos RM Osteoarticular EESSResonancia RedSalud ElquiAún no hay calificaciones
- Artefactos en UltrasonidoDocumento9 páginasArtefactos en UltrasonidoZulin K. TorresAún no hay calificaciones
- Aporte IndividualDocumento17 páginasAporte IndividualDayajimeAún no hay calificaciones
- Resonancia MagneticaDocumento49 páginasResonancia Magneticaroy_conejo6884Aún no hay calificaciones
- Técnicas Radiológicas II - Estudios Contrastados - Tracto Digestivo - Aparatoo UrinarioDocumento62 páginasTécnicas Radiológicas II - Estudios Contrastados - Tracto Digestivo - Aparatoo UrinarioWalter Solis ContrerasAún no hay calificaciones
- Anatomia de La ProstataDocumento34 páginasAnatomia de La ProstataChristopher Fernandez AlarconAún no hay calificaciones
- Resonancia MagneticaDocumento41 páginasResonancia MagneticaDavid Campos AvilaAún no hay calificaciones
- Resonancia DR HombroDocumento17 páginasResonancia DR Hombrosigrid100% (1)
- Resumen Tac Licenciatura 1-HardwareDocumento7 páginasResumen Tac Licenciatura 1-HardwareLeoTabaresAún no hay calificaciones
- Transito IntestinalDocumento6 páginasTransito IntestinalOrlando Solano SosaAún no hay calificaciones
- RD #000160-2021-Dg-Insnsb Guia de Resonancia MagneticaDocumento78 páginasRD #000160-2021-Dg-Insnsb Guia de Resonancia MagneticaErnesto García100% (1)
- Resonancia MagnéticaDocumento14 páginasResonancia MagnéticaRusbely MontillaAún no hay calificaciones
- Resonancia Grupo 1Documento13 páginasResonancia Grupo 1Josselyn Cruz100% (1)
- RADIOLOGIA DIGITAL PricipiosDocumento33 páginasRADIOLOGIA DIGITAL PricipiosElena CóndorazañeroAún no hay calificaciones
- Tomografía de Silla TurcaDocumento5 páginasTomografía de Silla TurcaRick Androll Aguirre0% (1)
- Efecto TalónDocumento12 páginasEfecto TalónAndrea Medina100% (1)
- Protocolodecerebrormn 150226174107 Conversion Gate01Documento38 páginasProtocolodecerebrormn 150226174107 Conversion Gate01joguyAún no hay calificaciones
- Radiación DispersaDocumento24 páginasRadiación DispersaYasser Farouk Sevilla HernandezAún no hay calificaciones
- Protocolos de TóraxDocumento10 páginasProtocolos de TóraxÁngelaAún no hay calificaciones
- Fist Ulog Rafi ADocumento8 páginasFist Ulog Rafi ACuauhtémoc JiménezAún no hay calificaciones
- HISTEROSALPINGOGRAFIA JPDocumento26 páginasHISTEROSALPINGOGRAFIA JPAstrid Pereira50% (2)
- Saturación Grasa y PrepulsosDocumento1 páginaSaturación Grasa y PrepulsosWilliams Oskar Estrada YbarraAún no hay calificaciones
- Tac Fisica Helicoidal 2010Documento31 páginasTac Fisica Helicoidal 2010Anonymous faRVVGNAún no hay calificaciones
- Introducción A La RadiologíaDocumento40 páginasIntroducción A La Radiologíarpalaciosf92% (12)
- Principios Fisicos de La Resonancia Magnetica Original PDFDocumento19 páginasPrincipios Fisicos de La Resonancia Magnetica Original PDFMiraldaperezAún no hay calificaciones
- Resonancia ManeticaDocumento14 páginasResonancia ManeticaDenisseIreneVillachicaAún no hay calificaciones
- Principios Fisicos Básicos de La Resonancia MagneticaDocumento5 páginasPrincipios Fisicos Básicos de La Resonancia MagneticaIsabel SantosAún no hay calificaciones
- La Física de La RMDocumento15 páginasLa Física de La RMMicaela AlvinoAún no hay calificaciones
- Curso de Resonancia Magnética Bioimagenes FAARDITDocumento9 páginasCurso de Resonancia Magnética Bioimagenes FAARDITCesar LopezAún no hay calificaciones
- Memoria Descriptiva GeneralDocumento4 páginasMemoria Descriptiva Generalosborn1Aún no hay calificaciones
- American Pitbull TerrierDocumento14 páginasAmerican Pitbull TerrierSebastianEnriqueMendozaPartidaAún no hay calificaciones
- Tesis de Escala ParentalDocumento33 páginasTesis de Escala ParentalMartha Elena Rodriguez Robles100% (2)
- Check List AutoelevadoresDocumento4 páginasCheck List AutoelevadoresBruno CamisciaAún no hay calificaciones
- Trabajo Precario y Salud MentalDocumento7 páginasTrabajo Precario y Salud MentalSilvinaCabrejasAún no hay calificaciones
- Practica 13 UREADocumento4 páginasPractica 13 UREALuis PachecoAún no hay calificaciones
- Practica #10 - Balanza de CorrienteDocumento10 páginasPractica #10 - Balanza de CorrienteMarco Antonio CuetoAún no hay calificaciones
- Efecto DopplerDocumento13 páginasEfecto DopplerAlejandro GuerreroAún no hay calificaciones
- Riesgos Ocupacionales en Odontología - Taller 2Documento6 páginasRiesgos Ocupacionales en Odontología - Taller 2Estefa BernaalAún no hay calificaciones
- Accesorios PARA ARENADORADocumento4 páginasAccesorios PARA ARENADORAAnonymous YqiomOTXAún no hay calificaciones
- Informe Exposición Procesos Industriales Tema EmbutidosDocumento18 páginasInforme Exposición Procesos Industriales Tema EmbutidosKevin Andre Fuentes100% (1)
- Sistema Circulatorio en Seres VivosDocumento5 páginasSistema Circulatorio en Seres VivossamuelarboledasAún no hay calificaciones
- Diseño Concreto A 700 KG/CM2Documento11 páginasDiseño Concreto A 700 KG/CM2Fernando Andree Salazar AlcantaraAún no hay calificaciones
- Bandura ExperimentoDocumento37 páginasBandura ExperimentoMorales Gonzales Francisco Abraham50% (2)
- Experiments Per Identificar PlàsticsDocumento9 páginasExperiments Per Identificar Plàsticsjdjgdjdjd456Aún no hay calificaciones
- Clonacion y TransgenesisDocumento38 páginasClonacion y TransgenesisMiguelViC100% (1)
- Unidad 2 Fase 3 Comercialización y Biotecnología Florícola-Colaborativo Abril 25-FinalDocumento11 páginasUnidad 2 Fase 3 Comercialización y Biotecnología Florícola-Colaborativo Abril 25-FinalArbey GonzalezAún no hay calificaciones
- Endemia, Epidemia y PandemiaDocumento4 páginasEndemia, Epidemia y Pandemiaroberto9020% (1)
- Sistema Endocrino y DigestivoDocumento13 páginasSistema Endocrino y DigestivoLuis DazaAún no hay calificaciones
- Butlleti Groc Nuevos HipoglicemiantesDocumento4 páginasButlleti Groc Nuevos HipoglicemiantesAgustina BazziAún no hay calificaciones
- Teoria Del Yin Fuego en Las Enfermedades Intricadas y DifícilesDocumento4 páginasTeoria Del Yin Fuego en Las Enfermedades Intricadas y DifícilesCABALLO66100% (3)
- Triptico MeridaDocumento2 páginasTriptico MeridayuliiAún no hay calificaciones
- Asfalto LiquidoDocumento7 páginasAsfalto LiquidoJairo PuertasAún no hay calificaciones
- Identificación de RocasDocumento7 páginasIdentificación de RocasYuliany SalazarAún no hay calificaciones
- ACTIVIDAD N°16 Plantas Sirven para Nuestra Alimentación SaludableDocumento6 páginasACTIVIDAD N°16 Plantas Sirven para Nuestra Alimentación SaludableAbigail Ashante TolentinoAún no hay calificaciones
- Consumo de Alcohol y Su Asociación Con El Rendimiento Academico.Documento7 páginasConsumo de Alcohol y Su Asociación Con El Rendimiento Academico.Omar Trejo hernandezAún no hay calificaciones
- Equipo OpDocumento5 páginasEquipo OpJose OlguinAún no hay calificaciones
- CatalogofcDocumento11 páginasCatalogofcAlejandro HernandezAún no hay calificaciones
- Infográfia Lactato 2Documento1 páginaInfográfia Lactato 2Angel Lopez LariosAún no hay calificaciones