También podría gustarte
- Glutamina en Nutricion ClinicaDocumento9 páginasGlutamina en Nutricion ClinicaGustavo Eduardo Canul MedinaAún no hay calificaciones
- Crisis HipertensivaDocumento41 páginasCrisis HipertensivaZCRYSTHELL100% (1)
- Liquidos BiologicosDocumento33 páginasLiquidos BiologicoscarlaAún no hay calificaciones
- Trastorno Con Déficit Atencional (TDA) : Docente Guia: Dra. Julieta Anllo. Integrantes: Constanza OrtegaDocumento28 páginasTrastorno Con Déficit Atencional (TDA) : Docente Guia: Dra. Julieta Anllo. Integrantes: Constanza OrtegaEsteban Pinto50% (2)
- 15 Clase de Perio Absceso Periodontal PDFDocumento5 páginas15 Clase de Perio Absceso Periodontal PDFLorenzo PanesAún no hay calificaciones
- Anestesia Con Bajos FlujosDocumento6 páginasAnestesia Con Bajos FlujosGiorceAún no hay calificaciones
- Drogas Anestésicas - ClasificaciónDocumento1 páginaDrogas Anestésicas - ClasificaciónGiorceAún no hay calificaciones
- Cómo y Cuando Revertir Los Relajantes MuscularesDocumento5 páginasCómo y Cuando Revertir Los Relajantes MuscularesGiorceAún no hay calificaciones
- Anestesia en El Paciente SépticoDocumento9 páginasAnestesia en El Paciente SépticoGiorceAún no hay calificaciones
- AROMA FileManager - El Primer Gestor de Archivos para El RecoveryDocumento5 páginasAROMA FileManager - El Primer Gestor de Archivos para El RecoveryGiorceAún no hay calificaciones
- Xposed Los Mejores Módulos para Xposed Framework - AndroidPITDocumento2 páginasXposed Los Mejores Módulos para Xposed Framework - AndroidPITGiorceAún no hay calificaciones
- Wanam Xposed - El Androide LibreDocumento4 páginasWanam Xposed - El Androide LibreGiorceAún no hay calificaciones
- Manual Del Propietario Dremel 3000Documento23 páginasManual Del Propietario Dremel 3000GiorceAún no hay calificaciones
- Xposed - Qué Es y Cómo Instalar PDFDocumento5 páginasXposed - Qué Es y Cómo Instalar PDFGiorceAún no hay calificaciones
- Códigos en NeumáticosDocumento7 páginasCódigos en NeumáticosGiorceAún no hay calificaciones
- Tutorial - Cómo Usar Correctamente El Titanium BackupDocumento7 páginasTutorial - Cómo Usar Correctamente El Titanium BackupGiorceAún no hay calificaciones
- Resetear Orinoquia Huawei Ascend Y221, Hard ResetDocumento2 páginasResetear Orinoquia Huawei Ascend Y221, Hard ResetGiorce50% (2)
- Tutorial Todo Sobre ClockWorkMod Recovery y WipesDocumento13 páginasTutorial Todo Sobre ClockWorkMod Recovery y WipesGiorceAún no hay calificaciones
- Recuperación Ante Un Softbrick o Un Bootloop Samsung Galaxy S3 (GT-I9300)Documento12 páginasRecuperación Ante Un Softbrick o Un Bootloop Samsung Galaxy S3 (GT-I9300)GiorceAún no hay calificaciones
- Cómo Saltarse El Patrón de Seguridad de Android - El Androide Libre PDFDocumento5 páginasCómo Saltarse El Patrón de Seguridad de Android - El Androide Libre PDFGiorceAún no hay calificaciones
- Deodexar ROM Stock, para Lus Usuarios de Samsung Que No Pueden Instalar Xposed - TuAndroidDocumento10 páginasDeodexar ROM Stock, para Lus Usuarios de Samsung Que No Pueden Instalar Xposed - TuAndroidGiorceAún no hay calificaciones
- Deodexar ROM Stock, para Lus Usuarios de Samsung Que No Pueden Instalar Xposed - TuAndroidDocumento10 páginasDeodexar ROM Stock, para Lus Usuarios de Samsung Que No Pueden Instalar Xposed - TuAndroidGiorceAún no hay calificaciones
- Cómo Instalar Kernels en Android de Manera SencillaDocumento3 páginasCómo Instalar Kernels en Android de Manera SencillaGiorceAún no hay calificaciones
- Cómo Saltarse El Patrón de Seguridad de Android - El Androide Libre PDFDocumento5 páginasCómo Saltarse El Patrón de Seguridad de Android - El Androide Libre PDFGiorceAún no hay calificaciones
- Manual de Neumologia Clinica 2edi PDFDocumento337 páginasManual de Neumologia Clinica 2edi PDFZonny SpellAún no hay calificaciones
- Informe Sobre Hipertension ArterialDocumento5 páginasInforme Sobre Hipertension ArterialJoshua MaciasAún no hay calificaciones
- Examen VirusDocumento8 páginasExamen VirusNorawaylandAún no hay calificaciones
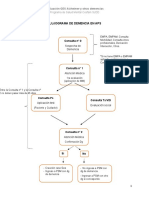
- Flujograma DemenciaDocumento1 páginaFlujograma DemenciaCristobal UrreaAún no hay calificaciones
- Hemorragias2 141110103212 Conversion Gate01Documento23 páginasHemorragias2 141110103212 Conversion Gate01Edwin Tapias100% (1)
- Arco de ErickDocumento6 páginasArco de ErickRiccy Lobo RodriguezAún no hay calificaciones
- Chavez Valdivia - Epidemiología - Casos y Controles y CohorteDocumento5 páginasChavez Valdivia - Epidemiología - Casos y Controles y CohorteJajayra Chavez ValdiviaAún no hay calificaciones
- ALBUM Sistema EndocrinoDocumento13 páginasALBUM Sistema Endocrinodantevela100% (2)
- Hemograma: Estudio Método: Impedancia Electrica + ColorimetriDocumento2 páginasHemograma: Estudio Método: Impedancia Electrica + ColorimetriVictoria achavalAún no hay calificaciones
- Leccion 5 QuemadurasDocumento10 páginasLeccion 5 QuemadurasOnaney J Herrand FerminAún no hay calificaciones
- Apuntes Examen Clinica de PequenosDocumento137 páginasApuntes Examen Clinica de PequenosFelix SanchezAún no hay calificaciones
- Guia para El Manejo R.P.BDocumento40 páginasGuia para El Manejo R.P.BEdgar Carreon CoronaAún no hay calificaciones
- Cuestionario Unidad Medicina NuclearDocumento3 páginasCuestionario Unidad Medicina NuclearFrancisco López del AngelAún no hay calificaciones
- Traumas de La Columna.Documento10 páginasTraumas de La Columna.melissa franco mosqueraAún no hay calificaciones
- Apuntes Trauma - TECDocumento6 páginasApuntes Trauma - TECDarelys LopezAún no hay calificaciones
- Enfermería PediátricaDocumento61 páginasEnfermería PediátricaDavid ChinchillaAún no hay calificaciones
- Ameu PasosDocumento1 páginaAmeu PasosCarlos BeltranAún no hay calificaciones
- Oximetazolina NebulizacionesDocumento4 páginasOximetazolina NebulizacionesAlexis MelgarAún no hay calificaciones
- Ictericia Neonatal 2Documento29 páginasIctericia Neonatal 2DARLIN CAMILA ENRRIQUEZ TERRAZASAún no hay calificaciones
- Examen Medico Completo - Edgar Alvarez PDFDocumento12 páginasExamen Medico Completo - Edgar Alvarez PDFsandra171717Aún no hay calificaciones
- Tesis Rosario UcvDocumento21 páginasTesis Rosario UcvAlma Yepez GamarraAún no hay calificaciones
- Ravos XDocumento11 páginasRavos XTeffy CoralAún no hay calificaciones
- Farmaco 123Documento3 páginasFarmaco 123Jesús David VergaraAún no hay calificaciones
- Cáncer de Colon Gastroenterologia RosaDocumento12 páginasCáncer de Colon Gastroenterologia RosaCoral PeñaAún no hay calificaciones
- Diapositivas LinfohematopoyeticoDocumento45 páginasDiapositivas LinfohematopoyeticoEnrique Luis Felix Lopez LabanAún no hay calificaciones