También podría gustarte
- Orden Poales.Documento5 páginasOrden Poales.Daga Gt AnAún no hay calificaciones
- 1-Poales TyphaceaeDocumento6 páginas1-Poales TyphaceaeyohanAún no hay calificaciones
- 2 BromeliaceaeDocumento9 páginas2 Bromeliaceaealfer688Aún no hay calificaciones
- Obtener Archivo RecursoDocumento30 páginasObtener Archivo RecursoBERTHA LUCIA MEDINA BECERRAAún no hay calificaciones
- Morfologia de La HojaDocumento73 páginasMorfologia de La Hojajdiaz23a4078Aún no hay calificaciones
- POACEAS ExomorfologiaDocumento29 páginasPOACEAS ExomorfologiaAriel BarrosoAún no hay calificaciones
- 1 FabaceaeDocumento20 páginas1 FabaceaeHMSAAún no hay calificaciones
- Marantaceae 97Documento7 páginasMarantaceae 97Juan David Rodriguez HurtadoAún no hay calificaciones
- División Magnoliophyta: M. Cs. Víctor Raúl Arévalo SaucedoDocumento26 páginasDivisión Magnoliophyta: M. Cs. Víctor Raúl Arévalo SaucedoJHOEL LLANOS ALVARADOAún no hay calificaciones
- 1-Alismatales + AraceaeDocumento10 páginas1-Alismatales + AraceaeyohanAún no hay calificaciones
- NyctaginaceaeDocumento5 páginasNyctaginaceaeMilady PocohuancaAún no hay calificaciones
- Familia CactaceaeDocumento8 páginasFamilia CactaceaeBelen GrandaAún no hay calificaciones
- 1-Alismatales + AraceaeDocumento10 páginas1-Alismatales + AraceaeDesiderioBernalAún no hay calificaciones
- ChenopodiaceaeDocumento3 páginasChenopodiaceaenelly carranza rodriguezAún no hay calificaciones
- Familia Geraniaceae Bot ExpoDocumento9 páginasFamilia Geraniaceae Bot ExpoJane100% (1)
- Solanaceae PDFDocumento11 páginasSolanaceae PDFtxakurlegiAún no hay calificaciones
- Flor 2016Documento24 páginasFlor 2016Eduardo YCAún no hay calificaciones
- División MagnoliophytaDocumento40 páginasDivisión MagnoliophytaJOSE DAVID IRIGOIN CAMPOSAún no hay calificaciones
- MonocotledoneasDocumento57 páginasMonocotledoneasalicia cardozoAún no hay calificaciones
- EuphorbiaceaeDocumento10 páginasEuphorbiaceaeMiguel Cordova BautistaAún no hay calificaciones
- Morfologia de Gramineas y LeguminosasDocumento15 páginasMorfologia de Gramineas y LeguminosasMvz Alfredo72% (18)
- Musaceae-EuphorbiaceaeDocumento21 páginasMusaceae-EuphorbiaceaeJose Luis Chasqui VelascoAún no hay calificaciones
- 4 CyperaceaeDocumento6 páginas4 CyperaceaejulianaAún no hay calificaciones
- Reino MetafitasDocumento13 páginasReino MetafitasSantiagoCamiloRiveraGomezAún no hay calificaciones
- 4 PlantaginaceaeDocumento7 páginas4 PlantaginaceaeNijan DereAún no hay calificaciones
- 6 ZingiberaceaeDocumento5 páginas6 ZingiberaceaejulianaAún no hay calificaciones
- HelechosDocumento15 páginasHelechosLeonardo Oziel TorresAún no hay calificaciones
- Clases de Pastos y Forrajes 2020 II - PA IIDocumento8 páginasClases de Pastos y Forrajes 2020 II - PA IIChinguel Calle HéctorAún no hay calificaciones
- Botan Sistem 4Documento95 páginasBotan Sistem 4ESTEFANY JACKELINE RABANAL BAZANAún no hay calificaciones
- Flora Del TolimaDocumento17 páginasFlora Del TolimaMiguel Angel Pertuz CordobaAún no hay calificaciones
- FabaceaeDocumento16 páginasFabaceaeMiguel Cordova BautistaAún no hay calificaciones
- Botánica Sistemática. Subdivisión GymnospermaeDocumento5 páginasBotánica Sistemática. Subdivisión GymnospermaeJoel RinconAún no hay calificaciones
- Botánica Magistral 4Documento10 páginasBotánica Magistral 4Ángel DejuanaAún no hay calificaciones
- BoraginaceaeDocumento4 páginasBoraginaceaearroyo_18davidAún no hay calificaciones
- Herbario DigitalDocumento7 páginasHerbario DigitalOwer JuradoAún no hay calificaciones
- Primero t3 PlantasDocumento14 páginasPrimero t3 Plantastelleznorvin07Aún no hay calificaciones
- SepaloideanosDocumento19 páginasSepaloideanosFedericoAún no hay calificaciones
- Ferriol López - Familia Compositae (Asteraceae) - Caracteres Generales.Documento7 páginasFerriol López - Familia Compositae (Asteraceae) - Caracteres Generales.Karina MirandaAún no hay calificaciones
- Hipertextos 2019Documento153 páginasHipertextos 2019Benjamin AlvarezAún no hay calificaciones
- Copia de LIBRO Clase 9 - Malvales A MyrtalesDocumento16 páginasCopia de LIBRO Clase 9 - Malvales A Myrtalesaimasolar2509Aún no hay calificaciones
- 3 EphedraceaeDocumento3 páginas3 EphedraceaeErlan RHAún no hay calificaciones
- 6 Definicion de SemillasDocumento29 páginas6 Definicion de SemillasYAMILA CASTILLOAún no hay calificaciones
- Unidad 3 Pradera y ForrajeDocumento22 páginasUnidad 3 Pradera y ForrajeDavid Martin SykoraAún no hay calificaciones
- Orden 31.rhamnalessDocumento5 páginasOrden 31.rhamnalessAnonymous 4r7KRWDYyh0% (1)
- 1 Asparagales OrchidaceaeDocumento14 páginas1 Asparagales OrchidaceaeyohanAún no hay calificaciones
- Familia CyperaceaeDocumento5 páginasFamilia CyperaceaeDANILOJITALA100% (1)
- CLASE 9 BotIIDocumento40 páginasCLASE 9 BotIIBelén AlarcónAún no hay calificaciones
- 4 AlismataleceaeDocumento4 páginas4 AlismataleceaeLAURA MARGARITA GRANADA ACOSTAAún no hay calificaciones
- Pastos y ForrajesDocumento102 páginasPastos y ForrajesHerlan Campos50% (2)
- Resumen Andinos IIDocumento31 páginasResumen Andinos IICésar CruzAún no hay calificaciones
- INFORME 7 - Descripción Morfológica y Especies Más Representativas de La Región Euphorbiaceae y PassifloraceaeDocumento14 páginasINFORME 7 - Descripción Morfológica y Especies Más Representativas de La Región Euphorbiaceae y PassifloraceaeTomas Rodas VargasAún no hay calificaciones
- Guia de Identificacion de - HeliconiaDocumento33 páginasGuia de Identificacion de - Heliconiaaristegl50% (2)
- 3 AmaryllidaceaeDocumento5 páginas3 AmaryllidaceaePaúl NúñezAún no hay calificaciones
- QuiscoDocumento42 páginasQuiscoMilagrosAún no hay calificaciones
- SolanaceaeDocumento2 páginasSolanaceaemaxAún no hay calificaciones
- Plantas Vasculares AngiospermasDocumento33 páginasPlantas Vasculares AngiospermasDiana PereiraAún no hay calificaciones
- Calorimetria (Termodinamica)Documento5 páginasCalorimetria (Termodinamica)Helen FabiolaAún no hay calificaciones
- 4 CannaceaeDocumento5 páginas4 CannaceaeHelen FabiolaAún no hay calificaciones
- Mehoramiento Del AlfalfaDocumento4 páginasMehoramiento Del AlfalfaHelen FabiolaAún no hay calificaciones
- ¿Que Es Lo Que HeredamosDocumento1 página¿Que Es Lo Que HeredamosHelen FabiolaAún no hay calificaciones
- ¿Que Es Lo Que HeredamosDocumento1 página¿Que Es Lo Que HeredamosHelen FabiolaAún no hay calificaciones
- Cabeza Tagma 1Documento15 páginasCabeza Tagma 1Helen FabiolaAún no hay calificaciones
- Tesis Especialista MinuDocumento66 páginasTesis Especialista MinuzinckortAún no hay calificaciones
- Huerto Urbano 12Documento44 páginasHuerto Urbano 12Katerin FAún no hay calificaciones
- Guia de Cuidado Planta CentavitoDocumento1 páginaGuia de Cuidado Planta CentavitodagobertoAún no hay calificaciones
- Plan de Cultivo - Almizcle 2021Documento12 páginasPlan de Cultivo - Almizcle 2021Omar CastellanosAún no hay calificaciones
- Los Beneficios de Aprender JardineríaDocumento4 páginasLos Beneficios de Aprender JardineríacarolinaAún no hay calificaciones
- CUESRIONARIO #1 Practica MecanizacionDocumento13 páginasCUESRIONARIO #1 Practica MecanizacionHerWil HTAún no hay calificaciones
- Gramíneas PoaceasDocumento44 páginasGramíneas PoaceasMaría MaryAún no hay calificaciones
- Tercera Practica Calificada Ec-511h-2020-1Documento2 páginasTercera Practica Calificada Ec-511h-2020-1Jordi ReyesAún no hay calificaciones
- fLORES Y FRUTOS PDFDocumento3 páginasfLORES Y FRUTOS PDFOsmar ValdezAún no hay calificaciones
- 1 Semillas - Cilantro - Patimorado - 250gDocumento4 páginas1 Semillas - Cilantro - Patimorado - 250gNancy yepesAún no hay calificaciones
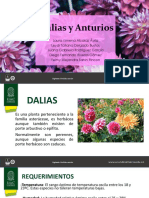
- Dalias y AnturiosDocumento14 páginasDalias y AnturiosValentinaAún no hay calificaciones
- Cuestionario #3Documento5 páginasCuestionario #3Abel Mauricio HuamánAún no hay calificaciones
- Este Es Un Encabezado de PáginaDocumento3 páginasEste Es Un Encabezado de Páginadaniel bernalAún no hay calificaciones
- Elemento Faltante - InformeDocumento13 páginasElemento Faltante - InformeSERGIO DARWIN RAMIREZ LINARES0% (1)
- Calculadora - Absorciones - Ex - Ante - v4 - tcm30-178912 (Autoguardado)Documento23 páginasCalculadora - Absorciones - Ex - Ante - v4 - tcm30-178912 (Autoguardado)pedroAún no hay calificaciones
- Tabla EquivalenciasDocumento4 páginasTabla Equivalenciasfabrizio reyes100% (1)
- HerramientasDocumento4 páginasHerramientasEva CalvoAún no hay calificaciones
- Proyecto Interdisciplinario .Julio MoraDocumento5 páginasProyecto Interdisciplinario .Julio MoraJulio hernan Mora floresAún no hay calificaciones
- Marco Teorico 5 y 6Documento3 páginasMarco Teorico 5 y 6Luis Eduardo VázquezAún no hay calificaciones
- La Tierra Como Material de ConstrucciónDocumento12 páginasLa Tierra Como Material de ConstrucciónJose De los herosAún no hay calificaciones
- Elementos de SilviculturaDocumento6 páginasElementos de SilviculturaAndres TelloAún no hay calificaciones
- Ciencia, Salud y Medio Ambiente, 2.° Grado - Semana 5Documento13 páginasCiencia, Salud y Medio Ambiente, 2.° Grado - Semana 5Carlos Armando CamposAún no hay calificaciones
- Asistencia TécnicaDocumento4 páginasAsistencia TécnicaEdward Castro UrrutiaAún no hay calificaciones
- 001 Panorama Grupal InteresesDocumento2 páginas001 Panorama Grupal InteresesJosefa Guzmán EspinosaAún no hay calificaciones
- Manual Basico para Hacer CompostDocumento8 páginasManual Basico para Hacer CompostFrancisca Bascuñán ArriolaAún no hay calificaciones
- Nuevas Alternativas Mi Natura Digital C01-2023 Al 02.01Documento4 páginasNuevas Alternativas Mi Natura Digital C01-2023 Al 02.01Lilly PinedoAún no hay calificaciones
- Catálogo #6 Bandejas de GerminaciónDocumento13 páginasCatálogo #6 Bandejas de GerminaciónGhkkk100% (1)
- Calculo Presion Constante en TuberiaDocumento3 páginasCalculo Presion Constante en Tuberiadulcemarielcarrasco7Aún no hay calificaciones
- Pro-Ato - 007 Procedimiento para Roceria Manual y MecanicaDocumento5 páginasPro-Ato - 007 Procedimiento para Roceria Manual y MecanicaKaren Delgado100% (1)
- Cancionero JarochoDocumento4 páginasCancionero JarochoSalvador OsnayaAún no hay calificaciones